Fizikai Szemle honlap |
Tartalomjegyzék |
Fizikai Szemle 1996/6. 195.o.
Lábos Elemér
SOTE, I. sz. Anatómiai Intézet, Budapest
Az elektrofiziológia (EF) nem azonos a neurofiziológiával vagy az ingerfiziológiával, de átfedésük jelentős. Minden sejt ingerlékeny. Válaszol az elektromos, kémiai és más ingerekre. Sőt egyen- (DC) vagy váltakozó (AC) áramot, illetve feszültséget is fenntart. Hagyományosan és pontatlanul különbséget tesznek ingerlékeny és nemingerlékeny sejtek között. Előbbiek az ideg- és izomsejtek: De egvsejtűeknek (Paramaecium), növényeknek (Mimosa, Nitella; Chara; gyökerek; fotoszintetizáló levél) pete-, vér-, hám- és mirigysejteknek is vannak passzív és aktív elektromos tulajdonságai és reagálnak ingerekre. Sőt sejtmagok belseje és külseje között is mértek potenciálkülönbséget (ecetmuslicák nyálmirigysejtjeiben). Ideg- és izomsejteknél tipikus esetben az elektromos feszültségkülönbség a sejt belseje és külseje közt alakul ki és ingerlésre legtöbbször gyorsan lefutó impulzus (spike = tüske) keletkezik és terjed, majd többnyire kémiai közvetítő anyagokkal újabb elektromos jelet vált más sejteken. Jellegzetes a külső ingerben és a válaszban foglalt energia aránya (trigger = ravasz-jelenség) is. A megfelelő inger energiája kicsi a válaszokban foglalt energiához képest.
A bioelektromosság töltéshordozói (+ és -) ionok és nem elektronok. Kritikus szerepet játszik a feszültségek fenntartásában a sejtek szinte molekuláris határhártyája. Sem az elektronokra, sem a sejt belsejének szerepére utaló direkt hipotézisek nem bizonyultak tartósnak, vagy az erre utaló állítások csak sajátosan igazak. Hosszú idő telt el, amíg az ideg- és izomaktivitást az elektromossággal kapcsolatba hozták. Megelőzően inkább más analógiákat kerestek. Így a francia felvilágosodás szerzői (Diderot, D'Alembert, Holbach), mechanikus (folyadék, pneuma) vagy éppen obskurus magyarázatokat adtak. Newton az idegekben és a látás során az éter" vibrációs mozgásairól beszél. Ez a korszak még a vitalizmusé volt. De ír az elasztikus és elektromos spiritus" mozgásáról is (Principia, cit. Brazier, 174). A statikus elektromosságot hatása még hiába jelezte, hogy az élő szövetek fogékonyak az elektromosságra (Gray). Pedig embereket tudtak elektromossá" tenni.
Ez az áttekintés az utolsó 200-300 évre nézve sem lesz teljes és az újabb kivételektől eltekintve másodlagos forrásokon alapul. Ez a prioritások megítélésében lehet zavaró. Nem feltétlenül igaz minden esetre minden állításunk.
Galvani
Egyesek - túlzással - de azt állítják, hogy az elektromosság és a bioelektromosság feltárásának története párhuzamos. Igaz, régóta ismertek az elektromos halak (sajgató halak; Arisztotelész, Plinius, Plutarkhosz); torpedószerű-fajok görög, egyiptomi ábrázolásokon, vagy a borostyán (elektron) dörzsölésével kapcsolatos egyes jelenségek (görögök, Gilbert), de a halak ütésének elektromos természetét nem állították. Nem ismerték a halak elektromosságának két kategóriáját, az erős áramütést és az érzékelő funkciót ellátó gyenge áramokat.
Mai ismereteink szerint körülbelül hat halcsalád (Torpedinidae, Mormyridae, Malapteruridae, Gymnarchidae, Gymnotidae, Asroscopus fajok), és körülbelül 200 halfaj használ támadásra vagy kommunikációra elektromos impulzusokat. Az ütőelektromosság 1-100 V, 1-10 A nagyságrendbe esik. Az igen finom (1
![]() V) elektromos érzékelő szerv, a Lorenz-ampulla cápáknál fordul elő. Az elektromos kommunikáció gyenge jelei ritmikus, 1-10 V közötti és csomagokban jelentkező 100-400 Hz frekvenciájú jelsorozatok. A támadó és érzékelő impulzustermelés nem zárja ki egymást. Az elektromos szervek sorba kapcsolt izom- és olykor mirigysejtek.
V) elektromos érzékelő szerv, a Lorenz-ampulla cápáknál fordul elő. Az elektromos kommunikáció gyenge jelei ritmikus, 1-10 V közötti és csomagokban jelentkező 100-400 Hz frekvenciájú jelsorozatok. A támadó és érzékelő impulzustermelés nem zárja ki egymást. Az elektromos szervek sorba kapcsolt izom- és olykor mirigysejtek.
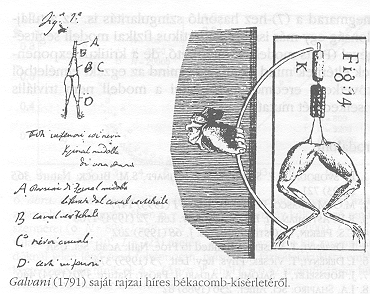
A régiek - in statu nascendi - elnevezéseikkel élve (a) dörzsvillamosságot (sztatikus elektromosság; leideni-palack) Kleist, 1745; Musschenbroek, Nollet, 1746), (b) érintkezési (galván, Galvani, Volta) villamosságot és (c) indukciós villamosságot (Faraday, 1831) különböztettek meg. Az elektroterápia - például szélhűdés gyógyítására - Galvani előtt a leideni palackkal kezdődött. (Boissier de la Croix de Sauvages, 1746). Mateuccinak tulajdonítják (1847) annak kimutatását, hogy az elektromos halak ütése" szikrát képes kiváltani. Így a nagyfeszültségű állati elektromosság létezését közvetlenül bizonyította. Előtte 50 évvel Cavendish modellkísérletei alapján már meggyőzte - legalábbis saját magát - arról, hogy az elektromos halak ütései elektromos természetűek. Tehát jelentősek ugyan az elektromosságtan és az állati elektromosság vizsgálatában a közös utak, de az első intenzív korszak Galvanitól átnyúlik a 19. századra, az elektrofiziológia megszületésének a századára.
Az (olasz) kortársak Fontanának tulajdonították az állati elektromosság létezésének feltevését (Caldini-iskola, 1750-1760). Nevezetes kísérletét Galvani 1791-ben írta le. A fémekkel érintkező békaizom ráng, összehúzódik. Volta ezt a fém-folyadék, illetve két eltérő fém (később galváncsipesz) és folyadék által létrehozott állandó feszültségnek tulajdonította. Azt is állította, hogy a galvánelem az elektromos természetű idegi folyadékot hozza mozgásba. Ezzel felfedezte a galvánelemeket, amelyeknek sok-sok válfaja már későbbi. Voltának igaza volt, de azt nem magyarázta meg, hogy miért hozhat létre külső egyenfeszültség izomrángást. Később Voltát Galvani-díjjal tüntették ki, amit Napoleon alapított (Brazier). Galvani viszont állását vesztette, amikor szembe került a napoleoni adminisztrációval. Galvani kísérlete példa a rosszul magyarázott, de korszakalkotó felfedezésekre. Volta szerint a békaizom az érintkezési elektromosságnak csak az akkor ismert legérzékenyebb indikátora. Galvani tudta azt is, hogy a békaizom vihar idején és leideni palack közelében is ráng, de ezt Caldini és Haller (német), már előbb említették.
Az állati elektromosság tényleges felfedezése
Du Bois Reymond (1849) mutatta ki, hogy a békaizom, vagy a békanyúltagy sértett fele negatív a sértetlenhez képest. Ő az elektrofiziológia megalapítója". A sértési (demarkációs) potenciál jó magyarázatát Hermannak (1897) is tulajdonítják, aki Du Bois Reymond, Pflüger és Vertigo magyarázatait hozta rendbe" elméletek alapján is (Overton, Nernst). Ringer (1880-1890) igazolta, hogy az izomrángáshoz a külső közeg megfelelő ionösszetétele kell. A sejteket hártya veszi körül, és a két oldalán kialakuló közeg potenciálkülönbsége észlelhető. Ma már sok potenciálmeghatározó folyamatot ismerünk (koncentrációkülönbség, ionmobilitás, diffúzió, szemipermeabilitás). Ezeket Nernst (1888-89) formulákba beleépítette: E = K ln (a1 / a2), ahol K= RT / nF, a1 és a2 (külső-belső) ionaktivitások. Később a Goldman-Hodgkin-Katz és rokonformulákban a koncentrációk; illetve aktivitások helyett azoknak permeabilitásokkal" súlyozott összege vagy más áll. Sok hasonló jó képlet létezik. Ezek mögött is a statisztikus mechanika (Boltzmann) és empirikus korrekciók állnak. A permeabilitások és arányaik nyugalomban és működés alatt eltérnek. A század végére Bernstein (1868-1902) szintézise (a membránelmélet) szerint a sejt belseje és külseje elektrolit, a kettő között hártya van, köztük izgalomkor megszűnő, sőt megforduló potenciálkülönbség áll fenn. Nyugalomban az ionfluxusok kicsik vagy elenyészőek. Az állítások később finomodtak. Bernstein a potenciál megfordulásáról szóló helyes állítását később módosította. Újból felfedezték.
A működési elektromosság (AP - action potential) 1819-ben hozta létre Oersted a tűs galvanométert. 1825-ben Nobili kimutatta, hogy a működő izom áramot termel. Bernstein, majd főleg Du Bois Reymond a működés elektromos jeleit már igen sok állati és emberi szövetben is észlelte. Matteucci (1845) egyik izommal mint áramforrással ingerelt egy másik izmot. Ugyanezt e század 30-as éveiben Arvanitaki idegekkel is kimutatta és ephapsis néven az érintkezést mesterséges szinapszisnak tekintette. A természetes elektromos szinapszisokat később fedezték fel. Az ingerlékeny szöveteket akár mechanikusan akár kémiai úton is lehet aktiválni, amikor az általa produkált impulzus (vagy hullám) közvetlenül vagy kémiai közvetítéssel átjut elsősorban a szinapszisoknak (Sherrington, Gray, Eccles) nevezett helyeken a vele érintkező másikra (ingerületáttevődés). Du Bois Reymond szerint a terjedő ingerület negatív" elektromos hullám. Ő választotta szét véglegesen a bioelektromosság két fundamentális formáját, a nyugalmi és az akciós áramot. Nála áll össze először az ingerlési (induktor) és az elvezetési technika (galvanométer). Azonban, az hogy a terjedés összehasonlíthatatlanul lassúbb, mint a fényé (szemben Müller állításával), már Helmholtz (1853) eredménye (körülbelül 1 mm/s - 100 m/s). Kísérlete ma is tanítható.
Az elektrotónus Du Bois Reymond elnevezése, idegekről és ideg-izom készítményekről van szó. Sejtek, szövetek felületén egyenáram hatására potenciálprofil alakul ki. Tőle származik az ingerlés, az elvezetés, analektro-, katelektrotónus kifejezések. Az itt a kérdés, hogy az ingeri.ilet elektromos hulláma milyen elektródnál keletkezik, milyen polaritású, hogyan váltható ki, vagy gyengíthető. Később kifinomult rángási szabályok alakultak ki (Pflüger, Waller), amelyeket a klinikumban is alkalmaztak. Egy didaktikailag és történetileg fontos jelenségcsoportról van szó. Magyarázatukhoz az elektródok elrendezése, száma lényeges, és a helyileg elvezetett, illetve a tovaterjedő ingerület fogalmai szükségesek.
Válaszok elektromos és adekvát ingerekre
Minden élő sejt így vagy úgy ingerelhető, és az ingerre (így vagy úgy) válaszol. Általában valamilyen ingerre kiemelkedő érzékenység és specificitás áll fenn. Ez az inger az illető receptor adekvát ingere. Az elektromos ingerlésre csaknem minden sejt, vagy sejtrészlet válaszol. De elektromosan nem ingerlékeny membránrészletekről is beszéltek (Grundfest) és ezt vitatták is (Kuffler). A specializált érzékenység szélsőséges példája fotoreceptorok egy-foton-érzékenysége (minden kvantum számít", Sakitt). Ezt pszichofiziológiai kísérletben emberen, Hecht igazolta, ma elektrofiziológiailag állati sejteken (Baylor et al.) is alátámasztották. Számítás ma is kell.
Az SDR korszak
SDR = strength-duration relationship. Mielőtt még jól mérhető amplitúdójú és időtartamú elektromos ingereket tudtak volna alkalmazni, már észrevették, hogy erősebb inger (áram, feszültség) után a válasz (izomrángás, idegimpulzus) korábban jelenik meg. Az összefüggést kezdetben formálisan hiperbolákhoz hasonlították. Vagyis az ingerküszöb (Bowditsch, 1871) csökken, ha nő az ingerelés időtartama. Ma jelentősége visszaszorult. Pedig ez az összefüggés vezetett a szöveteket elektromosan helyettesíteni vélt áramkörök konstrukciójához (Lapicque, Rushton, Blair, Hill, Monnier, Rashewsky, Solandt, 1900-1938, és mások). Tanulmányozásuk vezetett az akkomodáció felfedezéséhez is. Ez az elektromos ingerküszöb ingerlés hatására történő növekedése.
Ekvivalens áramkörök
Az élő - különösen az ingerlékeny - szövetek reagálnak az elektromos ingerekre és termelnek" is elektromosságot, azaz tartalmaznak telepet. Felmerült, hogy van-e ellenállásúk, vezetőképességük, kapacitásuk, induktivitásuk; azaz lineáris áramköri tulajdonságaik? Reagálnak-e ezek DC vagy AC hatásokra? Fokozatosan kiderült, hogy e tulajdonságok kiterjeszthetők más élő szövetekre, sejtekre, sejthártyára és változhatnak is a külső vagy belső tényezők által kiváltott ingerület" során. A mai ekvivalens áramkörök az SDR-elemzésekből és a Kelvin-típusú kábelek analógiájából származtathatók.
Kiderült az is, hogy csak nem-lineáris áramkörök írják le jól az elektromos ingerületi hullámokat. A lineárisak (például LItt + RIt + (1/C) I = Vt ) csak küszöb alatt használhatók úgy-ahogy és egyes elemek speciális problémákat vetnek fel. Mérések szerint a sejthártya kapacitása ingerület alatt alig változik vagy állandó marad, valódi induktivitások létezése pedig igen vitás. Az ellenállások feszültségfüggőek (nem ohmikusak). A technikai áramkörökkel szemben az áramköri paraméterek (R, C, L; U) nem koncentrálódnak, hanem elosztottak. Ennek a dinamikai leírásban következményei vannak: nemcsak közönséges (ODE), hanem parciális differenciálegyenletek (PDE) is szükségesek.
Bernstein (1902) a század elején Nernst (1888-89) munkájára alapozva az élő sejteket koncentrációs elemeknek tekintette. A sejt belseje jól vezető elektrolit-oldat, ami egy vékony, csaknem ionimpermeábilis hártyával van körülvéve. Nyugalomban a belső és külső fázis között potenciálkülönbség (a belső negatívabb) ellenállás és kapacitás van. Aktivitás alatt ez a nyugalmi membránpotenciál (Erest; MP) eléri a nullát (rövidzár). Ma tudjuk, hogy az előjel jelentősen meg is fordul. Ez a változás az akciós potenciál (AP). Kohlrausch (1876) már észlelte, hogy az élő szövetek ellenállása" nagyobb mint a halott szöveteké, és mint a testfolyadékoké. Ezt Fricke (1923-1925) kísérletesen alátámasztotta. Az "élő" vérszuszpenziók vezetőképessége csökken, ha a sejtsűrűség nő (Stewart, 1899).
A sejtellenállás" nem ohmikus, és valódi impedancia van. A hipotetikus áramkör alapján becsülhetők az R, L és C lineáris paraméterek. A kapacitás becslése az időállandóknak és aktiváló áram frekvenciafüggésének a mérésén alapul. A linearitás a nyugalmi potenciál kis környezetében fennáll, de távolabb már fellép a vezetőképesség feszültségfüggése és rektifikáció. Valójában impedanciát, reaktanciát, admittanciát mérhetünk és vizsgálhatjuk a mérőáram frekvenciájától és erősségétől való függést. Negatív ellenállásról is jogosult beszélni.
A sejthártya kapacitása vagy szuszpenzióban, vagy egyetlen sejten határozható meg. Ha például tudnánk a nyugalmi ellenállást és egy négysvögimpulzusra adott integrált hullám alakú válasz időállandóját, akkor a kapacitás kiszámítható. Nagyságrendje 0,5 - 10
![]() F/cm2. Onnan ered, hogy két vezető közeget elválaszt a vékony (lipid kettősrétegből + beágyazott fehérjékből álló) szigetelő" réteg. A kapacitív áram It = CVt. A kapacitás maga rétegvastagságtól, felülettől, permittivitástól, polarizálhatóságtól függ. A membrán itt szereplő vastagsága 300-1000 nm. A kapacitás viselkedésének az ioncsatornák kapuzó áramainak mérésében is szerepe van. A membránellenállást és kapacitást párhuzamosan kapcsolva szokták leírni.
F/cm2. Onnan ered, hogy két vezető közeget elválaszt a vékony (lipid kettősrétegből + beágyazott fehérjékből álló) szigetelő" réteg. A kapacitív áram It = CVt. A kapacitás maga rétegvastagságtól, felülettől, permittivitástól, polarizálhatóságtól függ. A membrán itt szereplő vastagsága 300-1000 nm. A kapacitás viselkedésének az ioncsatornák kapuzó áramainak mérésében is szerepe van. A membránellenállást és kapacitást párhuzamosan kapcsolva szokták leírni.
A tintahal óriás axonjának a longitudinális és transzverzális induktivitását Cole és Baker a harmincas években meghatározták. A mérés érdekes vitát váltott ki. Egyesek szerint az induktív tag formális és nem mond semmit a geneziséről, mások szerint nem-lineáris egyenletekre a lineáris kategóriák (RCL) nem igazán alkalmazhatók.
Lineáris áramkörökben az ohmikus ellenállás állandó. De élő szövetekben inger hatására már Benedict vizsgálatai alapján a múlt században következtettek a csökkenésére. A sejthártya ellenállásának ingerületet kísérő, feszültségfüggő megváltozásai Hodgkin és Huxley (HH) vizsgálatai óta tudatosodtak, bár az ingerületben lévő axon vezetőképességének fokozódását Cole és Curtis (1938-39) mutatta ki. Azaz az ingerlékeny szövetek ellenállása nem ohmikus.
A fontosabb ionokra nézve (Na, K, Cl, Ca) Nernst-formulák alapján úgynevezett egyedi egyensúlyi potenciálokat számíthatunk, feltéve, ha ismerjük a belső és külső koncentrációkat (aktivitásokat). Az RT/F faktor értéke körülbelül 54-62. Ha az extracelluláris referenciaelektród potenciálja zérus, akkor tipikus potenciálok az alábbiak: EK = -98 mV, ECl = -90 mV Emembrán = -90 mV, ENa = +67 mV, ECa = +129 mV (emlős vázizomrostra). Ha viszont küszöb alatti potenciált kémiai ágens vált ki, akkor van olyan DC előpolarizáció, ahol ez előjelet vált (megfordulási potenciál). Ebből szoktak a töltéshordozóra következtetni. Az izgató posztszinaptikus potenciál (depolarizációs, EPSP) megfordulási helye zérus mV felett, a gátlóé (hiperpolarizációs, IPSP) a K és Cl potenciál között van.
Az I- Vkarakterisztikák egyenestől (Ohm-törvény) való eltérése az egyenirányítás jelensége. Biológiai objektumokban legalább négy rektifikáció-típust lehet kimutatni: depolarizációs (inward) aktiváció, depolarizációs inaktiváció, hiperpolarizációs (outward) aktiváció és hiperpolarizációs inaktiváció. E változatokról és elnevezésekről (reguláris és anomális, azaz hiperpolarizációs aktivációs, illetve pillanatnyi és késleltetett) Grundfest ad áttekintést.
Módszerek
A kezdetleges galván és induktív ingerlés számos változatát dolgozták ki és e század első harmadában még használták is. Itt csak Duchenne és Du Bois Reymond szikrainduktoraira hivatkozunk. Ezek ingadozó áramütést adnak: sem az idő sem az intenzitás nem jól szabályozható. A galváncsipesz egyenáramú forrás és ez is bizonytalan. Az ugrást a fejlődésben a négyszöghullámok jelentették. Lehetővé vált továbbá a szinuszos, a lineárisan, exponenciálisan változó ingerimpulzusok alkalmazása: Megvalósítási generációi léteztek (elektroncsövestől az erősen integráltig).
A múlt században higanyos-kapilláris, majd a finomabb húros galvanométerekkel mérték a kis feszültség- és áramingadozásokat. E században alakult ki a fejlett erősítéstechnika. Igen korán úgynevezett elektrométererősítők, majd válogatott FET (field effect tranzisztoros) erősítők, végül ezek integrált generációi jelentkeztek. Ugyanis 1
![]() V -100 mV, illetve 1 pA -1
V -100 mV, illetve 1 pA -1
![]() A tartományokban kell mérni nem túl gyors jeleket (0-20 kHz). A jelforrások belső ellenállása meghaladhatja a
100 k
A tartományokban kell mérni nem túl gyors jeleket (0-20 kHz). A jelforrások belső ellenállása meghaladhatja a
100 k![]() -100 M
-100 M![]() -ot. Az igényes bemenő fokozatok mögött jó minőségű (tárolóernyős) oszcilloszkóp vagy ma már digitalizált számítógépes végső mérőrendszer áll. A mérési eredményeket még 30 éve FM magnetofonokon tárolták. A hardcopy készítése pedig fotokimográfokkal" történt. Ma már (szelekció és feldolgozás után) számítógépekről nyomtathatók. A mérés vizuális, sőt akusztikus követése ma is fontos, mert egy jó kísérlet nem vak, hanem interaktív, azaz kísérlet közben változtatni érdemes. Sokszor versenyt kell futni a preparátum életidejével is.
-ot. Az igényes bemenő fokozatok mögött jó minőségű (tárolóernyős) oszcilloszkóp vagy ma már digitalizált számítógépes végső mérőrendszer áll. A mérési eredményeket még 30 éve FM magnetofonokon tárolták. A hardcopy készítése pedig fotokimográfokkal" történt. Ma már (szelekció és feldolgozás után) számítógépekről nyomtathatók. A mérés vizuális, sőt akusztikus követése ma is fontos, mert egy jó kísérlet nem vak, hanem interaktív, azaz kísérlet közben változtatni érdemes. Sokszor versenyt kell futni a preparátum életidejével is.
A hagyományos áram-, feszültség- és patch-clamp eljárások mérőhidakat, kompenzációs lehetőségeket, visszacsatolásokat, műveleti erősítőket és egyéb sajátos technikákat is igényelnek.
Az elektródok az elektrofiziológiában ingerlésre és a keletkezett elektromos jel (lassú vagy impulzusszerű hullám) elvezetésére szolgálnak. De az anyagok bevitelére (iontoforézis), részletek kiiktatására ([jelző]lézió, szövetpusztulás okozására), anyagok specifikus észlelésére (pH mérésére, ionok, molekulák detektálására), vagy akár vezérlésre is használhatók. Régi csúcs a 0,06 mikron csúcsátmérőjű wolframelektród. Amit észlelnek, az nem mindig egy sejttől ered, sőt a klinikai célú EEG és EKG csak szöveti finomságú. Több vonalon futó irányzat a sok sejtet, forrást vagy generátort szeparáltan észlelni tudó elektródsokaság. Minden ingerléshez, illetve elvezetéshez két elektród szükséges. Ezek között lehet közös, de akkor gondos értelmezés is kell. A mikroelektródok legtöbbször üvegkapillárisok (Ling és Gerard) vagy hegyezett wolframhuzalok.
Elméleti háttér is kell. A bioelektromos jelenségekhez mindig kell és van is - jó vagy rossz - előfeltevés és háttérmagyarázat. Ez bármely kísérletben befolyásolhatja annak lefolytatását is. Kísérletes munkákban megengedik az eredmények taglalását". Itt lehet egy artefaktumot eredménynek minősíteni, leírást eltúlozni vagy rosszul magyarázni". Axióma az, hogy az eredményeket úgy kell elfogulatlanul" leírni, ahogyan (legjobb belátásunk szerint) azok történtek. Ez nehezebb mint gondolnánk. Az eredmények matematikai kiegészítése sokszor történik színvonalatlanul és gyakran megáll rosszul indikált, de jól elvégzett statisztikai próbáknál. Ezzel akár egy artefaktumot szintaktikusan ugyan, de igazolni is lehet.
Hodkin és Huxley (HH, HésH) egyenletek
1952-ben öt közlemény eredménye a mai napig domináns ingerületelmélet és változatai. HésH már 1939-1945 között először vezetett el tintahal óriás axonjából intracellulárisan. Kimutatták a nyugalmi potenciál megfordulását (lezárva ezzel a Bernstein shunt-hipotézis érvényességét). Hodgkin és Rushton 1946-ban a polarizálás hatására kialakuló potenciálprofilt lineáris kábelegyenlettel (PDE) írták le. A későbbi áttörésben a Cole és Marmont (1949) által kifejlesztett feszültségzár (voltage clamp) módszer játszott döntő szerepet. Akciót tudtak létrehozni akciós potenciálhullám nélkül két elektróddal. A második (az áram) elektród körében lehetett mérni a fixáláshoz szükséges áramot. A módszer nem mindig alkalmazható és metodikai problémák is felmerülnek. Megkülönböztetnek feszültség és áram-clampet, továbbá időbeli és térbeli fixálást. Az egyenletrendszernek - amelyek mindegyikét áramsűrűségekre írják fel - az alábbi változatai a fontosak: (1) - A teljes 4 változós forma 4 egyenlettel. (2) Az időbeli V-clamp-egyenlet, amelyben Vt = 0 (V = const). A vele arányos kapacitív áram zérus. A maradékáramot tekintik transzmembránális ionáramnak. (3) Áram-clamp vagy tér-clamp rendszer. Négy változó, 4 nem-lineáris egyenlet. Ezt vizsgálták leggyakrabban. (4) Utazóhullám-egyenlet, amelyik alakváltozás nélküli, állandó sebességű akciós hullámot tételez fel. Az egyszerű feltevések ellenére zavarbaejtően instabilis, nem reális. Öt változó (V, Vt, m, h, n), négy egyenlet. Az eredeti fő változók az alábbiak: V(x, t), m (x, t), h (x, t), n (x, t), amelyek rendre a potenciál, a Na+-aktiváció (m), Na+-inaktiváció (h), K+-aktiváció (n). Mindegyik egydimenziós térváltozótól és időtől függ. Az elméletben, sikerei dacára, problematikus az idő és tér-clamp szelektív tárgyalhatósága, a bonyolultabb sejtgeometria esetei és a ma már igen sok ioncsatorna figyelembe vétele.
Kapuk, pórusok, csatornák és vezérlésük
A bernsteini membrán-hipotézis kulcsfogalma a membrán áteresztőképessége. Nyugalomban globálisan jóval kisebb, mint ingerületkor vagy aktív gátlás alatt. Akció alatt áteresztő kapuknak kell szelektíve megnyílni, majd záródni a 10-20 nm méretű ionok (sőt más molekulák) számára. Az átjutás helyei a pórusok, a nyitás-zárás neve a kapuzás (ami töltésmozgással, azaz sajátos árammal is jár), a csatornák pedig egy (vagy olykor több) ion számára szolgáló speciális szelektív pórusok. Vezérlésük (nyitás, illetve zárás) membránfeszültséggel vagy speciális molekulákkal történik.
Az ionmozgásokkal járó áramoktól eltér a töltött kapuanyagok (például fehérjék) nyitáshoz-záráshoz vezető mozgását (konformációját) kísérő áram, a kapuáram. A fehérjemozgások pikosecundum töredékétől mikrosecundum tartományig terjednek, míg az akciós áramok millisecundum rendűek. Létezésüket HésH már 1952-ben megjósolta. Mérhetők, illetve számíthatók mérési anomáliákból", a sejthártyák feszültségválaszára adott kapacitív áramának aszimmetriája alapján. (Stämpli és mtsai).
Néhány csatornaféleség: (1) - Na+-csatornák. A leghomogénebb (V függő, TTX érzékeny) csoport, de mégsem egységes. (2) - K+-csatornák. - 2.1 V-függő, 2.2.1 K(Ca) kalciumaktivált nagy (BK) és 2.2.2 kis (SK) vezetőképességű-csatorna, 2.3 A K-spike-ok csatornái. (3) - Ca+++ csatornák. 3.1 Gyors az N és T típus és 3.2 lassú az L típus. (4) - Cl--csatornák. Különböző szövetekben és fajokban eltérő típusokat írnak le. Stabilizáló hatásuk van. (5) Multi-ion csatornák. Ezek a többnyire ligandum vezérelt csatornák (ACh, GABA, ATP). A csatornák jellemzése a töltéshordozóval, a dinamikával, a külső és belső gátló és aktiváló tényezőkkel, az előfordulási hellyel és sok egyébbel jellemezhető. Átfogó munka Hille monográfiája és hasznos az Annual Reviev of Physiology sok írása 1979-1985 között. Megjegyezendő még, hogy: (1) az úgynevezett ion-pumpa mechanizmusok lassú folyamatok, és (2) valódi ingerküszöb nem mérhető, de igen jó absztrakció. (3) továbbá kölcsönhatásokkal és komplikált dinamikájukkal számolni kell. (4) az elnevezések jelzői sokfélék és azonosításuk körültekintést igényel.
Az ionáteresztés szelektivitásában szerepet játszik az átmérőjük. Az ionmobilitás az ionok tömegétől is függ. Vizes oldatokban dinamikus vízburok veszi körül őket, ami átmérőjüket megkeveri. De a hidratált átmérő fogalmát Hille zavaros fogalomnak véli. A kolloidika és az elektrokémia sok szempont szerint rakta sorba az ionok egy-egy homogén csoportját. Eisenman azt vette észre és elméletileg is alátámasztotta, hogy például az alkáli fémionok 120 lehetséges sorrendje közül mindössze 11 fontos. Az átmérősorrendeknek ellentmondó változatok alapján kölcsönhatásokra következtetnek.
Szinapszisok és hálózatok
A szinapszis idegsejtek és/vagy végrehajtó sejtek olyan érintkezési felülete, ahol kémiai, illetve elektromos jelátadás történik. Fő részei" a preszinaptikus végződés, szinaptikus rés és posztszinaptikus sejthártyarészlet együttese. A fogalom Sherringtontól ered, aki sohasem látott szinapszist. Tipikus kémiai ingerületáttevődés az alábbi: a szinapszishoz érkező elektromos jel anyagfelszabadulást vált ki, az átkerül a résen és egy posztszinaptikus elektromos jelet vált ki. Ebbe a képbe ma már beletartoznak esetleg a G-fehérjék, a foszforilációs mechanizmusok, a másodlagos vivőanyagok (cAMP, cGMP) felszabadulása és a modulációk" is, több átvivő anyag, eltérések az elágazódások végén stb. Közeledünk a szubcelluláris elemzések lehetőségéhez. Elektrofiziológiájuk zöme Eccles felfedezéseivel indult. A szinapszisokkal kapcsolódó sejtek neuronhálózatokat képeznek.
Patch-clamp
A sejthártyán az ionok meglehetősen szelektív molekuláris járatokon, az ioncsatornákon át jutnak ki vagy be. Ma már hordozómolekulákat nem tételeznek fel. Ezt sokáig csak mint igen jó hipotézist támasztották alá a kapuáramokra utaló jelek, majd egy új technika, a patch-clamp igazolta. 1975 táján vezette be Neher-Sakmann-Steinbach. A módszerhez a régóta ismert membránpotenciálzajok és a miniatűr szinaptikus potenciálok elemzése (és a HH-V-clamp) vezettek (Neher).
A módszer abban áll, hogy a vizsgált sejthez egy igen nagy shunt-ellenállással
(G![]() ) illeszkedő üvegmikroelektródot szívnak". Így 1-10 pA csatornaáramok és 1-100 pS vezetőképességek válnak mérhetővé. Négy formája van: (1) a sejt felületének egy darabját izolálják el, (2) a sejtmembrán meg is nyílik, (3) a sejtmembrán felszívott darabja (patch) ki is szakad és a sejtbelső vagy (4) a sejtkülső tapad az üvegelektród csúcsához. Így a sejthártya kis darabjának lokális (mikro)viselkedését, azaz izolált csatorná(ka)t dinamikusan lehet vizsgálni. A jellegzetes regisztrátum két vagy több domináns amplitudószint között ingadozó áramokat mutat. Ma már nemcsak nagy sejtek, hanem 1-100 négyzetmikronnyi hártyadarab vizsgálata is lehetséges. A külső és belső fázis közötti feszültség fixálható, ami azért fontos, mert a sejtmembránok ioncsatornáinak vezetőképessége általában feszültségfüggő. A négy technika kissé eltérő vizsgálatokat tesz lehetővé. Helyszelektív anyaghatás elemzéseket, csatornák eloszlását sejtfelületeken (direkt mozaik, illetve heterogenitás vizsgálat).
) illeszkedő üvegmikroelektródot szívnak". Így 1-10 pA csatornaáramok és 1-100 pS vezetőképességek válnak mérhetővé. Négy formája van: (1) a sejt felületének egy darabját izolálják el, (2) a sejtmembrán meg is nyílik, (3) a sejtmembrán felszívott darabja (patch) ki is szakad és a sejtbelső vagy (4) a sejtkülső tapad az üvegelektród csúcsához. Így a sejthártya kis darabjának lokális (mikro)viselkedését, azaz izolált csatorná(ka)t dinamikusan lehet vizsgálni. A jellegzetes regisztrátum két vagy több domináns amplitudószint között ingadozó áramokat mutat. Ma már nemcsak nagy sejtek, hanem 1-100 négyzetmikronnyi hártyadarab vizsgálata is lehetséges. A külső és belső fázis közötti feszültség fixálható, ami azért fontos, mert a sejtmembránok ioncsatornáinak vezetőképessége általában feszültségfüggő. A négy technika kissé eltérő vizsgálatokat tesz lehetővé. Helyszelektív anyaghatás elemzéseket, csatornák eloszlását sejtfelületeken (direkt mozaik, illetve heterogenitás vizsgálat).
Soksejt és extracelluláris egység-regisztrátumok
Az EKG, EEG, kiváltott potenciálok és egyéb makroelektródos eljárások sok sejt együttes tevékenységét regisztrálják egy vagy sok elektródával. De sokszor a fémmikroelektródokon extracelluláris elvezetésekkor is több forrásból jövő impulzusok láthatók. Felmerül több probléma: (1) - Hogyan lesz az idegsejtek impulzusaiból két külső elektród között mérhető potenciál? (Lorente de No térfogati vezetése, Rosenfalck analízise). (2) - Hogyan lehet a tömegregisztrátumokat forrásokra és nyelőkre" felbontani? (Freeman-Stone, 1969 - Mitzdorf-Singer, 1975). A soksejt-elvezetések matematikája kifinomult, de nem mindenható. (3) - Amplitúdó és frekvencia spektrumanalízis. (4) Igen sikeresek a direkt inger-válasz elemzések (Hubel, Wiesel).
Elektroaktív festékek
Mintegy 20-25 éves metodikai fejlemény a bioelektromos tevékenységre optikai változásokkal reagáló anyagok, az elektroaktív festékek használata. A festék fényelnyelése, fényemissziója, kettőstörése, dikroizmusa változhat. Az MP változásának ez a mérése szinte nem invazív, szemben az intracelluláris üveg és más beszúrt elektródokkal. Kis sejtek sokaságának aktivitása vizsgálható vele. Vannak lassú és gyors festékek. Közöltek már 1020000-nyi regisztrációs pont 1 ms-nál kisebb időfelbontású méréséről is adatokat (emlős hippocampus szeletek, más kérgi területek esetében), A hasznos impulzust a zajból sokszor, de nem mindig jelátlagolással emelik ki. Nem feltétlenül egysejt-elvezetésekről van szó. Előfordul sejtek részeinek vagy sejtek sokaságának együttes tevékenységét közlő munka is. E módszer sem abszolút ártalmatlan. A tér-időfelbontás, illetve jelerősség által felvetett problémákon túl a festékek elhalványodása, farmakológiai és a fotodinámiás hatásuk (károsító fotooxidáció) korlátokat szabhatnak. Ismert festékek a lucifer sárga, a merocianin származékok. Hátránya még a módszernek az is, hogy felszíni sejtek vagy agyszeletek vizsgálatára kell szorítkozni.
Nem esett szó mindenről
(1) Például az elektroenkefalográfia (EEG) történetéről, pedig klinikai jelentősége ma is van. 1929-37 között Berger több közlésben terjesztette. Kihagytuk az EMG (myo-), EOG (olfacto-), ERG (retino-) és más makropotenciálokat is.
(2) Nem esett szó a villamos áram és villámcsapás káros hatásairól, a villamosszékről, az elektrosokkról mint tüneti terápiás eljárásról.
(3) Az ingerületvezetés kapcsán nem tértünk ki a szaltatórikus vezetés, a magvezetés és a lokális mikroáramok tényeire, illetve elméleteire.
(4) Hanyagoltuk az éber állatokon végzett vagy a tanulás mechanizmusaival közvetlenül összefüggő elektrofiziológiát...
(5) Nem tárgyaltuk a hálózatok elektrofiziológiai elemzésének lehetőségeit.
(6) Léteznek anomális, de biztosan nem általánosan előforduló elektromos jelek. Ilyenek a retina sok sejtjének fordított polaritása vagy a spike-ok nélküli ingerületáttevődés. De nem tipikusak az elektromos szinapszisok sem.
(7) Vannak abnormális elektromos jelek is.
(8) Az óriásaxonokon alkalmazható perfúzió módszere a külső és a belső ionmilieu tetszőleges változtatását tette lehetővé.
(9) Speciális anyagok optikai tulajdonságaikkal reagálnak a kalciumkoncentráció megváltozására.
(10) Mennyiségek vagy jelenségek méréséhez, illetve kimutatásához sajátos kísérleti paradigmák", műszeres és kémiai ellátmány", továbbá a kiértékelést megengedő elméleti-matematikai formalizmus is tartozik. De a részletezés e munkát lehetetlenül felduzzasztaná.
Az artefaktumok kimeríthetetlenek
Itt az artefaktum is egy olyan faktum, amit nem az a tényező (faktor) hoz létre, amelynek esetleg tulajdonítható lenne. Például egyenfeszültségek mérésekor olyan nem biológiai elektródpotenciálok" hatnak, amelyek a folyadék-fém határrétegnek vagy üvegfelületnek tulajdoníthatók. Ingerléskor pedig olyan jelszerű impulzusok lépnek fel, amelyeket elhalt szövet elektromos sajátságai okozhatnak. Rokon kategóriák az eljárások invazív" jellege folytán észlelt hatások. Durva elektród csökkenti a membránpotenciál nagyságát (az értékek nőttek is a technika javulásával). Az anyaghatások és anyagkimutatások esetében az álpozitív és álnegatív elnevezések a szokványosak: vagyis ott is tündért látunk, ahol nincs, vagy ott sem észleljük, ahol van. Az elektrofiziológus nevezetes artefaktuma az eszközök bemenetére árnyékolás és földelés" nélkül bejutó 50-60 Hz. Különösen akkor makacs, ha igen kis jelet kell észlelni és nagy erősítéseket használnak. Ezért e frekvenciatartomány biológiai eredetének megítélése körültekintést igényel. Hangkijelzésnél megszólalhat valamilyen rádióadó is, ami szűréssel kiiktatható.
A perspektívák sokasága
A történelem végéről beszélni Fukuyama óta megalapozatlan divat. Neves magyar neurofiziológus vetette fel előttem azt, hogy a patch-clamp-pel az elektrofiziológia kimerítette önmagát. Szerintem az elektrofiziológia legfeljebb átmenetileg stagnál vagy lokálisan visszafejlődik.
Magyarországon gyakran a nem kielégítő színvonal, a tapasztalatlanság és olykor a szakmai dilettantizmus társul a rossz anyagi lehetőségekkel. Az elektrofiziológia iskoláját végig kell és lehet (egyetemen) járni. Doktrínákat megcáfolni és finomabb (például molekulaközeli") kísérleteket végezni csak képzett emberek képesek és csak csúcstechnikával. Másrészt az elektrofiziológiai módszerek bármelyikének igen nagy a gyógyszeripari jelentősége is.
Irodalom
G. ADELMAN (ed ): Encyclopedia of Neurosciences - Birkhauser, Basel. 1987.
TH.H. BULLOCK, G.A. HORRIDGE: Structure and Function in the Nervous Systems of Invertebrates - Freeman et Co., San Francisco 1965.
J. BURES, M. PETRAN, J. ZACHAR: Electrophysiological Methods in Bíologicall Research - CzASc Publ. House, Prague 1960, 1967.
K.S. COLE: Membranes, Ions and Impulses - Univ. of California Press, Berkeley, 1968.
DUBOIS REYMOND (1841-1849): Untersuchungen über thierische Elektrizität - Berlin, Reiner.
J.C. ECCLES: The Physiology of Synapses - Springer, Berlin 1964.
L. GALVANI: (1791): De Viribus Electricitatis in Motu Musculari - Reprint: Licht, Cambridge, (transl, by R.M. Green, 1953)
H. GRUNDFEST: Comparative Electrobiology of Excitable Membranes - Adv. Comp. Physiol. and Biochem. 2 (1966) 2-116.
L. GOTTSCHICK: Die Leistungen des Nervensystem - Fischer, Leipzig, 1955.
HELMHOLTZ: Vorlaufiger Bericht uber die Fortpflanzunggeschwindigkeit der Nervenreiung - Arch. Anat. Physiol. 85 (1850) 71-73
A.V. HILL: Excitation and accomodation in nerve - Proc. Roy. Soc. B, 119 (1936) 305-355.
B. HILLE: Ionic Channels and Excitable Membranes - Sinauer Ass., Sunderland, Mass. 1992.
A.L. HODGKIN, A.F. HUXLEY: A quantitative description of membrane current and its application to conduction and excitation of in nerve - J. Physiol. 117(1952) 500-544.
J.A. STAMFORD (ed.): Monitoring Neuronal Activity. A Practical Approach - IRL, Oxford UP, Oxford, PA-Series, 1991, 294 p.
F.G. WORDEN, J.P. SWAZEY, G. ADELMAN: The Neurosciences: Paths of Discovery - MIT Press, Cambridge, Mass. 1975.