Fizikai Szemle honlap |
Tartalomjegyzék |
Fizikai Szemle 1997/1. 21.o.
Varjú Dezső
Biokibernetikai Tanszék,
Tübingeni Egyetem, Németország
A második világháború idején Norbert Wiener amerikai matematikust arra buzdították, hogy töprengjen el azon, miként is lehetne megjavítani a légelhárító ágyúk célbatalálását. A feladat egy matematikai probléma: előzetes megfigyelések, mérések alapján gyorsan ki kell számítani, hol lesz a repülőgép, amikor a véges sebességgel haladó lövedéknek célba kellene érnie. Egyenesen röpülő bombázó esetén ehhez elég ismerni a gép és a lövedék sebességét; a kettő viszonyának megfelelően kell a repülőgép elé lőni. Vadászgépek esetén súlyosabb a probléma, mert azok irányokat gyorsan tudják változtatni. Nem tudom, lett-e Wiener munkájának valami gyakorlati haszna is, de a célbatalálás problémája őt - úgy látszik - nagyon lenyűgözte, mert a háború után is sokáig foglalkoztatta. Nem technikai okokból, hanem azon felismerés miatt, hogy a célbatalálás az élőlények számára is egy örök feladat.
Az élőlények célirányos mozgása
Úton lévő állatok többnyire nem céltalanul bolyonganak, hanem egy jól meghatározott helyre akarnak eljutni, valahova, ahol például táplálékot, védett helyet, előnyösebb időjárási viszonyokat vagy egy társat találhatnak.
Mondjuk egy gnúcsorda tagjai akkor is némi harapnivalóra, lelhetnek, ha véletlenszerűen bolyonganak a szavannában, de nagy bajban lennének, ha ivóvízhez is ilymódon kellene jutniuk. Lakóhellyel rendelkező állatok rendszeresen elhagyják fészküket, odújukat, hogy élelmiszert gyűjtsenek, mint például a méhek vagy a fiókáikat etető madarak, de utána haza kell találniuk. A költöző madarak vagy számos más, szárazföldi és tengeri állat is több-ezer km-t vándorol, de a megfelelő évszakban visszatér a szülőhelyére.
Ezen esetekben a célok ugyan - a repülőgépekkel ellentétben - nem mozognak, de mozgó célokra is van számos példa az állatvilágban: hím legyek egyértelmű céllal repülnek a nőstények után; vadászó állatok üldözik áldozataikat, amelyek sokszor irányt változtatnak. De álló célok esetén is szükség van az irány többszöri vagy folyamatos javítására. A célba törekvő állatokat elsodorja a szél vagy a vízáram, el kell kerülniük az akadályokat, vagy pedig egyszerűen csak az érzékszerveik és az idegrendszerük fogyatékosságai miatt átmenetileg rossz irányba mozognak, míg aztán a hiba észlelhetővé nem válik. Wienernek az volt a nézete, hogy e tájékozódási feladatok kivitelezéséhez az idegrendszernek hasonló matematikai feladatokat kell megoldania, mint a légelhárítóknak. Úgy gondolta, hogy ez a problémakör egy új tudományággá válhatna, és az ógörög hajókormányos, a Kybernetes neve után kibernetikára keresztelte azt.
Az önműködő szabályozás
A kibernetika feladatköre idővel lényegesen kibővült. Ennek egyik oka az, hogy a célba való törekvés során egy olyan folyamat játszódik le, amit a mérnökök önműködő szabályozásnak neveznek. Pontosabb megjelölése: önműködő szabályozás negatív visszacsatolással. Az élőlényeknek valahogy érzékelniük kell, hogy a pillanatnyi útirány nem a célba vezet. Ha például egy bizonyos szöggel balra térnek el a helyes iránytól, akkor egy megfelelő nagyságú fordulatot kell tenniük jobbra.
Az ilyen negatív visszacsatolásos szabályozásoknak se szeri se száma az élővilágban. Az élet legfontosabb előfeltétele ugyanis az, hogy bizonyos fiziológiai paraméterek értéke állandó legyen, illetve csak szűk határok között mozogjon. A határok átlépése sokszor visszafordíthatatlan folyamatokat gerjeszt, ami halálhoz vezethet. A legismertebb példa talán a melegvérű állatok testhőmérsékletének a szabályozása. Ez ugyanazon elv alapján működik, mint a modern lakások hőszabályozója. Egy eszköz méri a pillanatnyi hőmérsékleti értéket. Ha az alacsonyabb, mint a kívánt hőmérséklet, akkor egy másik eszköz nyitja a fűtőtest csapját; fordított esetben zárja. Az élőlényeket az evolúció többnyire nem csak egy mérő és egy szabályozóeszközzel látta el. A test hőmérsékletét például érzéksejtek mérik a bőrben, valamint az agyban, a fül és az agyat vérrel ellátó nagyartériák közelében. Az utóbbi azért lényeges, mert egy állandó hőmérséklet fenntartása az idegsejtek számára sokkal fontosabb, mint például a végtagok számára. A számos szabályozó folyamat között ugyancsak jól ismert az izzadás túl magas és a didergés túl alacsony hőmérséklet esetén. A libabőr nálunk, embereknél egy egyáltalán nem hatékony maradványa az evolúciónak, mert a mi szőrzetünk már egészen visszafejlődött. Kevésbé közismert a bőr alatti finom érelágazások szerepe. Magasabb hőmérsékletek esetén ezek kitágulnak, és ezáltal elősegítik a törzsből odajutó túl meleg vér lehűlését, feltéve, hogy a környező levegő hőmérséklete alacsonyabb, mint a véré. Alacsonyabb testhőmérséklet mellett összeszorulnak az erek, és így csökkentik a hőveszteséget. E folyamatokon kívül nagy szerepet játszik a viselkedés is. Kánikula esetén a legostobább kutya is elvonul az árnyékba, mi pedig a környezet hőmérsékletének megfelelően öltözködünk. A nagyszámú, más, önműködően szabályozott paraméterek közül én itt csak a közismertebbeket említeném, mint például a vérnyomást, a pulzust, a vérben lévő különböző vegyületek - például cukor, zsír - koncentrációját.
Egy egészen más kategóriába tartozik például a test egyensúlyának szabályozása. Testünk egyensúlya fizikai okokból eléggé labilis, és aktív szabályozás hiányában előbb utóbb elterülnénk. A fent említett esetekkel ellentétben ezt a képességet gyermekként kell megtanulnunk. A tanulás után ez a szabályozás is önműködően játszódik le, és csak akkor veszünk tudomást róla, ha valami zavar lép föl, mint például, ha valaki becsíp, és ezért tántorog.
Kölcsönzés a mérnöktudományoktól
Az önműködő negatív visszacsatolásos szabályozás kvantitatív leírására már a század eleje óta léteztek matematikai módszerek a mérnöki tudományokon belül. Ennek ellenére - egy kivétellel - csak az ötvenes évek elején kezdtek el a biológusok azon töprengeni, hogy vajon lehetne-e e módszereket hasonló biológiai folyamatok leírására is használni. E biológusok egyike Erich von Holst volt. Nem tudom, ismerte-e Wiener ötletét, minden esetre volt egy mérnök ismerőse, és a vele való beszélgetések során meggyőződött arról, hogy e módszerek nagyon hasznosak lehetnének az említett célra. Egy 1958-ban Karlsruhében tartott ülésen úgy nyilatkozott, hogy e módszerek olyan fontosak a biológusok számára, hogy nekik kellett volna kidolgozniuk őket, ha a mérnökök nem bocsátották volna a rendelkezésükre. A gyűlésen, amelyen én is résztvettem, az volt a benyomásom, hogy Erich von Holstot kisebbrendűségi érzések gyötörték a fizikusokkal szemben, akik az élettelen világ törvényszerűségeit kezdettől fogva kvantitatívan, matematikailag írták le. Úgy vélte, hogy a fizikusok szemében a biológusok nem számítanak komoly természettudósoknak, hiszen ők csak rovarokat, növényeket gyűjtögetnek, és megpróbálják azokat osztályozgatni. Persze ez nem volt igaz, mert már a múlt század eleje óta számos biológus foglalkozott az inger és a reakció viszonyának egzakt feltárásával. Azt is felismerték, hogy az ingerek többsége olyan reakciót vált ki, amely - egy negatív visszacsatolásnak megfelelően - az inger hatását csökkenti. Fáradozásuk persze csak az összefüggések tapasztalati leírására korlátozódott.
A biokibernetikusok fő törekvése az volt, hogy ezeket az összefüggéseket matematikailag fogalmazzák meg, és így kitalálják, milyen számítási feladatokat old meg az idegrendszer és hogyan. A kísérletekben alkalmasan választott ingerektől kiváltott reakciókat mértek. Itt persze jogos egy csomó kritikus kérdés: Milyenek az alkalmasan választott ingerek? Mire jó az, ha a reakció és az inger közötti összefüggést matematikailag is megfogalmazzuk, azaz felállítunk egy matematikai modellt? Mennyire megbízhatók e modellek? Kizárólagosak-e, vagy elképzelhetők más változatok is? Két példa segítségével - melyeket számtalan lehetőség közül választottam ki - megpróbálok választ adni e kérdésekre.
Az emberi szem pupillájának fényreakciója
A pupilla (írisz) reakciójának szerepe az, hogy a lehetőségekhez mérten szabályozza a recehártya megvilágítását. Ha növekszik a környezetben a fényerősség, akkor beszűkül a pupilla, aminek következtében a recehártya megvilágítása csekélyebb mértékben növekszik, mint a környezet fényintenzitása. Ha a fényerősség csökken, akkor a pupilla kitágul. A folyamat tehát egy tipikus önműködő szabályozás negatív visszacsatolással. Hogy ebben az esetben melyek az alkalmas ingerek, azt szintén a mérnöki matematika, pontosabban a rendszerelmélet adja meg: a fényerősség hirtelen megváltozása, vagy egy villanófény, vagy pedig a fényerősség periodikus modulációja. Az ezen ingerek által kiváltott reakciókat matematikailag egyszerűen lehet megfogalmazni, legalábbis lineáris rendszerek esetében.
Az inger és a reakció közötti összefüggés matematikai leírására elvben elég lenne egyetlen ingertípus által okozott reakció mérése. A biztonság kedvéért mindhárom ingertípus által kiváltott reakciót szokás mérni. A matematikai modell nagyjából akkor megbízható, ha mindhárom esetben elég pontosan leírja az összefüggést. Így volt a pupilla esetében is. A dologban egyrészt az a fontos, hogy tudjuk milyen matematikai műveletet kell az idegrendszernek elvégeznie. Marad persze a kérdés, hogyan oldja meg az idegrendszer e feladatot. Erre később visszatérek.
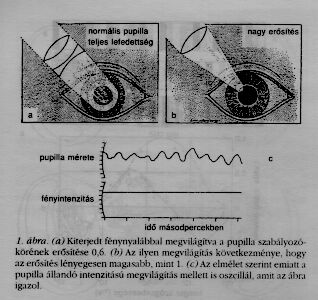
A másik fontos szempont, hogy a modell segítségével jósolni lehet a reakciót minden másfajta ingerre is. E jóslatok esetenként majdnem hihetetlenek, és ha kísérletileg igazolhatók, akkor egy további bizonyítékot szolgáltatnak arra, hogy a modell megfelel a valóságnak. Egy ilyen jóslat a pupilla esetében nagy port vert fel az akkori időkben. Az előző kísérletekben a szem egy olyan fénynyalábbal volt megvilágítva, melynek átmérője nagyobb volt, mint a pupilla maximális átmérője, és a nyaláb tengelye a pupilla közepén ment át (1.a ábra). A mérések eredménye szinuszosan modulált fényintenzitás esetén az volt, hogy a pupilla egy bizonyos frekvenciánál (körülbelül 1,5 Hz) 180 fokos fáziseltolódással reagált. A negatívan visszacsatolt szabályozó rendszerek elmélete szerint ilyen esetekben a rendszer elveszti stabilitását, ha a visszacsatoló szervek erősítése nagyobb, mint 1. Kiderült, hogy természetes körülmények között és az 1.a ábrán mutatott megvilágításnál is az erősítés csak 0,6, így a rendszer stabil.
A jóslat szerint a szabályozókör labilis lenne, és körülbelül 1,5 Hz-cel oszcillálna, ha sikerülne az erősítést 1 fölé növelni. De hogyan? Egy amerikai emberélettanos kollégám ötlete szerint ez elérhető, ha egy állandó intenzitású keskeny fénynyalábot úgy vetítenek a szemre, hogy az részben áthatoljon a pupillán, részben pedig az íriszre essék (1.b ábra). Utóbbi esetben a pupilla átmérőjének csekély változása erősen megváltoztatja a recehártya megvilágítását. A jóslat kísérletileg igazolódott (1.c ábra). A kísérlet könnyen megismételhető egy zseblámpa felhasználásával, melynek sugarát fókuszálni lehet.
A rovarok sebességérzékelése
A rovarok - több más állathoz hasonlóan - igyekeznek az optikai környezetük képét a recehártyájukon stabilizálni. Ha például a szél a levelet, amelyen egy bogár ül, balra fordítja, akkor az állat az így fellépő képeltolódást egy jobbra irányuló testforgással kompenzálja. A laboratóriumban persze nem a bogarat forgattuk, hanem az rögzítve volt egy henger közepében, melynek belső fala függőleges fekete és fehér csíkokkal volt kitapétázva (2.a ábra). A henger egyenletes sebességgel forgott a bogár körül, ami úgy volt felfüggesztve, hogy tudtunkra adhatta, milyen gyorsan fordulna, ha hagynánk (további részleteket lásd [1]-ben).
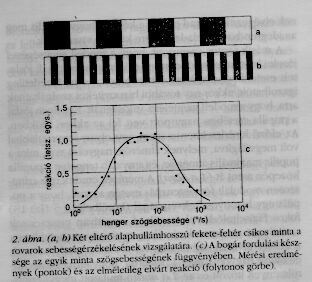
Három kísérleti eredmény különösen említésre méltó, ha az első kettő nem is nagyon meglepő. (1) A bogár fordulási készsége eleinte növekszik a henger szögsebességével, utána viszont csökken, és végül nulla lesz. Ez persze várható is volt, hiszen egy igen gyorsan forgó fekete-fehér minta számunkra is szürkének látszik, "elmosódik", úgyhogy mozgását nem tudjuk észlelni.(2) Az állat fordulási készsége a minta csíkjainak szélességétől függ. Ha egy kísérletben negyedére csökken a csíkszélesség (2. b ábra), akkor a rovar forgási tendenciája négyszer alacsonyabb szögsebességnél éri el a maximumát. Ez azt jelenti, hogy a bogarak tulajdonképpen nem a sebességet érzékelik, hanem azt, hogy milyen gyakoriak a fekete-fehér átmenetek egy érzősejt látóterében. (3) Egy kritikus érték alatti csíkszélesség esetén a bogár forgási tendenciája előjelet vált. A bogár nem úgy fordul tehát, hogy csökkentse a külvilághoz képesti szögsebességét, hanem ellenkező irányban.
Ezek és néhány további eredmény elegendőek voltak egy modell felállításához, amely a bogár fordulási készségét a henger szögsebességének függvényében kitűnően szimulálta (2.c ábra). Hangsúlyozom, itt nem arról van szó, hogy a mérési eredményekhez valamilyen alkalmas matematikai függvényt illesztettünk, amely esetleg csak éppen ezen eredményt írná le. Itt egy olyan modellről van szó, melynek segítségével a reakció bármely más fajtájú inger esetén is megjósolható. Az egyik ilyen kísérletileg igazolt jóslat volt például a reakció függése a henger szögsebességétől egy olyan henger esetén, melynek felülete szabálytalanul elhelyezett fehér, különböző árnyalatú szürke és fekete csíkokkal volt borítva.
A leglátványosabb eredmény azonban a következő volt: A fenti, két, csíkos mintát először úgy szuperponáltuk, hogy fekete + fekete = fekete, fehér + fehér = fehér, és fekete + fehér = szürke lett (3.a ábra). A második esetben ezen új mintában a nyíllal jelölt fekete és fehér csíkok fel lettek cserélve (3.b ábra). Végül a keskenysávú minta el lett tolva egy fél csíkszélességgel a szélessávúval szemben, és a két mintát ezután a fenti szabályoknak megfelelően szuperponáltuk (3.c ábra). Meg szeretném jegyezni, hogy az emberi szem számára az a és a b minta közötti különbség alig tűnik fel, míg a b és c minták különbözőnek látszanak. Hogy mit jósol a modell, azt a 3.d ábra görbéi mutatják, az utólag elvégzett mérések eredményeit pedig a különböző jelek. Az a mintára a jóslat szerint a bogár fordulási készsége úgy függ a henger szögsebességétől, mint az egyszeri, fekete-fehér csíkok esetén. Meglepő volt a jóslat a másik két mintával kapcsolatban egyrészt azért, mert a reakció előjelet vált a szögsebesség függvényében: alacsony sebességeknél negatív, és csak magasabb sebességeknél lesz pozitív. Másrészt pedig azért, mert a számunkra igen különbözőnek tűnő b és c minták azonos reakciót váltanak ki, míg a számunkra alig megkülönböztethető a és b minták nagyon különbözőek a bogár számára.
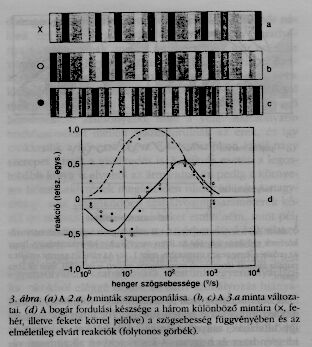
E jóslatok kísérleti igazolása volt az egyik legszebb sikerélményem. Mint a pupilla esetében, ezek az eredmények is arra utalnak, hogy a modell megbízhatóan enged azon műveletekre következtetni, melyeket az idegrendszer végez. Hasonló példákat százával lehetne még felsorolni.
Neuroanatómia és elektrofiziológia
Visszatérek arra a kérdésre, hogyan végzi el az idegrendszer a modellek szerint neki tulajdonított műveleteket. A viselkedés alapján erre a kérdésre nem tudunk választ adni, hiszen az idegsejtek szintjén számos lehetőség van ugyanazon művelet megvalósítására. Az eljárást, melynek segítségével erre a kérdésre szeretnék választ adni, egy régi történet segítségével szemléltetem. A háború után a keleti országok technológiája nem tudott lépést tartani a Nyugattal. Az ötvenes évek elején Nyugaton piacra dobták az akkor legmodernebbnek számító Tektronix 500 jelzésű oszcilloszkópot. A Szovjetunóban beszereztek valahogyan két példányt ebből a mérőeszközből. Az egyiket érintetlenül hagyták, és olyan vizsgálatokat végeztek rajta, melyeket én a pupillával és a bogarakkal kapcsolatban vázoltam: A bemenet - az inger - az időben különféle módon változó feszültség volt, és vizsgálták a kimenetet - a reakciót -, vagyis a képernyőn megjelenő görbéket. A másik készüléket felnyitották. Először csak azt rajzolták le, hogyan vannak összekötve az alkatrészek, utána meg azt vizsgálták, hogy milyen feszültségek mérhetők a különböző csomópontokban. Ehhez feltehetően az első érintetlen eszközt használták. Később teljesen szétszedték a második készüléket, és egyenként vizsgálták az alkatrészek tulajdonságait. Így a végén majdnem olyan minőségű oszcilloszkópokat sikerült gyártaniuk, mint amelyen a két eredeti volt.
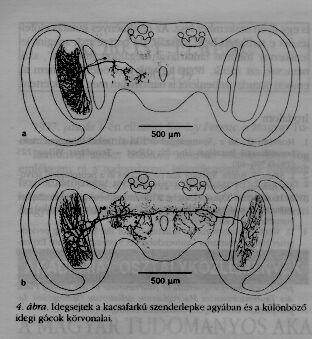
Hasonló az eljárás a biológiában is. Ha sikerült magatartási kísérletekben kitalálni, milyen műveleteket kell elvégeznie az idegrendszernek, és arra a kérdésre is választ szeretnénk kapni, hogyan végződnek el e műveletek, akkor be kell hatolni a szervezetbe. Az eljárás nagyon hasonló a keleti mérnökök módszeréhez. Manapság úgy lehet festeni egyedi idegsejteket az összes finom elágazásaikkal együtt, hogy ezek a mikroszkópban láthatóvá válnak (4.a, b ábrák). A sejteket egy doktoranduszom a kacsafarkú szenderlepke vizuális rendszerében festette meg.
Rovarok esetében ugyanazon sejtek azonos helyeken találhatók minden példány esetében. Festés nélkül az idegrendszer elég homogén szövet, és a mikroszkóp alatt nem lehet az egyes idegsejteket megkülönböztetni. Sok sejt festése után és megfelelő gyakorlattal meg lehet állapítani, milyen hálózatot alkotnak azok, hol történik a jelek továbbítása. A jelek elektromos impulzusok, melyeket finom elektródok segítségével le lehet vezetni, és az oszcilloszkóp ernyőjén láthatóvá tenni (5. ábra). A szóbanforgó esetben mindkét idegsejt inger nélkül is bizonyos számú impulzust gerjesztett másodpercenként. Inger esetén, annak tulajdonságától függően, az impulzusszám vagy növekszik, vagy csökken. Az ingerek fekete-fehér csíkos minták voltak, melyek a nyíllal jelzett irányban mozogtak. Az elektródok, melyek segítségével az impulzusokat levezetik, tölcséralakban kihegyezett, finom üvegcsövek. Hegyük átmérője csak néhány ezred milliméter, így nem okoznak nagy kárt, ha beleszúrják őket az idegsejtbe. Az elektródok elektromosságot vezető folyadékkal töltöttek, és egy vékony fém vezetőn át rá vannak kötve a mérőeszközökre (erősítők, oszcilloszkópok, személyi számítógép).

Az elektromos impulzusok levezetése és a sejt festése egy munkaütemben történik. Először az impulzusokat vezetik le, majd ugyanazon elektródon át a sejtet festékanyaggal töltik fel. Mindezen lehetőségeket - melyekről 40 évvel ezelőtt még csak álmodni sem tudtak a kutatók -az elektronika és a vegyipar hihetetlen gyorsaságú fejlődésének köszönhetjük. A jövő még sokkal többet ígér. A biológusok - többnyire mérnökök és fizikusok segítségével - olyan eljárások kifejlesztésén dolgoznak, melyek lehetővé teszik számos, egymással összekötött idegsejt aktivitásának egyidejű regisztrálását. Ilyen mérések segítségével sokkal gyorsabban meg lehet állapítani, miként kommunikálnak egymással a sejtek, hogyan oldják meg azokat a feladatokat, amelyeket a matematikai, kibernetikai modellek alapján elvárunk. A pillanatnyi eredmények még szerények, de sokat ígérőek.
A biokibernetika jövője
Végül néhány szó arról, hogyan látom én a biokibernetika jövőjét. Ehhez előbb vissza kell pillantanom a múltba. Mérnöknek vagy fizikusnak csak egy megfelelő egyetemi képzés után nevezheti magát valaki. A biokibernetika és biokibernetikus nem védett fogalmak, mindenki nevezhette és nevezheti magát így. Amikor az első, látványosabb eredmények ismertté váltak - a két fenti példa is közéjük tartozott - gombamódra nőttek a földből a "biokibernetikusok". A baj az volt, hogy sokan közülük el akarták hitetni a közönséggel - televízióban, rádióban, írott sajtóban -, hogy a biokibernetika egy csodaszer, melynek segítségével hamarosan meg fogjuk érteni, miként is működik az emberi agy, holott az akkori eredmények mai szemmel nézve igen szerények voltak, mint a példáim is, és csak egy speciális, elszigetelt funkcióját írták le az idegrendszernek.
A számítógépek fejlesztésének is nagy volt abban a szerepe, hogy az említett kollégák úgyszólván egy kalkulátorrá degradálták az agyat. Tették ezt azzal a mottóval, hogy "azt már értjük, miként működik a számológép, és mivel a lényeget tekintve az agy sem más, mint egy ilyen masina, annak a működését is hamarosan meg fogjuk érteni". Könyvek jelentek meg olyan címekkel, mint például "Mit gondol egy számológép?" vagy "Az elektromos agy", "Automaták és az ember". Így aztán a közönség nehezen tudott különbséget tenni a komoly tudomány és a fikció között. Mindenesetre fennállt a veszély, hogy aki biokibernetikusnak nevezi magát, azt esetleg eleve nem veszik komolyan. Ezért sokan, akik kezdetben biokibernetikusnak vallották magukat, máshogyan jelölték meg munkaterületüket, különösen az angol nyelvű országokban.
Werner Reichardt, a biokibernetika talán legismertebb úttörője Németországban volt olyan bátor, hogy a Max Planck Társaság általa alapított és vezetett kutatóintézetét
"Biokibernetikai Intézetnek" nevezze. Ezután én se tétováztam tovább, és tanszékemet, melynek eredeti neve Állattani Tanszék volt, szintén Biokibernetikai Tanszékké kereszteltem. Reichardt hozta létre 1961-ben a Springer Kiadóvállalatnál a Kyberraetik című tudományos folyóiratot, melynek neve 1975 óta Biological Cybernetics. A szerkesztőbizottság 25 tagja nemzetközileg ismert és elismert kutató. Ők gondoskodnak arról, hogy csak színvonalas közlemények jelenjenek meg a lapban. Következésképpen reputációja világszerte jó.
E tények ellenére kevés azon kutatóintézetek és egyetemi csoportok száma, melyek kutatási területüket biokibernetikának nevezik, holott ez az elnevezés sok esetben találó lenne. Sőt, a tendencia csökkenő annak ellenére, hogy a biokibernetikai módszerekkel elért eredmények az emberi agy működésének a megértésében is egyre jelentősebbek. A viszonylag egyszerű önműködő szabályozási és vezérlési folyamatok tanulmányozásán túl nagyon divatba jött manapság az úgynevezett kognitív neurobiológia. Reichardt utódja is e területen dolgozik, és a Tübingeni Egyetem Biológiai Karának döntése szerint az én utódom is egy ilyen szakember lesz. Az eredményes kutatás előfeltételei e területen nem rosszak, de persze már megint jelentkeztek túlfűtött fantáziával megáldott "tudósok", akik nemcsak azt ígérik, hogy agyunk működését, hanem az emberi öntudat mibenlétét is hamarosan meg fogjuk érteni.
Irodalom
_____________________
Előadás a Magyar Mérnökök és Építészek Svájci Egyesülete találkozásán (1996. szeptember 17.)