Fizikai Szemle honlap |
Tartalomjegyzék |
Pósfai Mihály
Pannon Egyetem, Veszprém
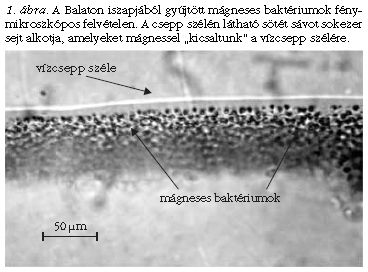
A vándormadarak, méhek, lazacok vagy angolnák bámulatos tájékozódása a Föld mágneses terében közismert. Kevesebben tudják, hogy egyes baktériumok is képesek a mágneses térhez igazodva navigálni, a számukra legkedvezőbb életteret megtalálni. Richard Blakemore [1] 1975-ben tavi mikroorganizmusokat vizsgált fénymikroszkóppal, és észrevette, hogy egy csapatnyi sejt a vízcseppnek mindig ugyanarra az oldalára úszott. Azt gyanította, hogy ezek fény felé úszó baktériumok, ezért elsötétítette a helyiséget. Mikor ez nem változtatott a baktériumok mozgásán, egyéb módosításokkal próbálkozott, ám hasztalan: a baktériumok továbbra is mindig a vízcsepp ugyanazon oldalára gyülekeztek. Ekkor egy furcsa ötlete támadt, és egy mágnest helyezett a csepp másik oldalára. A sejtek azonnal megfordultak, és a csepp túlsó oldalán gyűltek össze (1. ábra ). E felfedezés óta beszélünk magnetotaxisról, azaz a környezet mágneses terének irányához igazodó mozgásról, illetve magnetotaktikus (vagy egyszerűbben mágneses) baktériumokról.
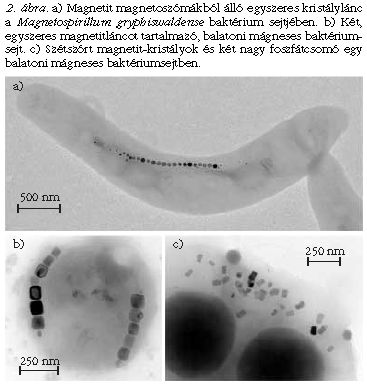
A mágneses baktériumok jellemzőtulajdonsága, hogy magnetoszómákat tartalmaznak (2. ábra). A magnetoszóma egy ferrimágneses nanokristályból (amely lehet Fe3O4 magnetit vagy Fe3S4 greigit) és az ezt körülvevő biológiai membránból áll [2]. A magnetoszómák rendszerint láncba vagy láncokba rendeződnek, de akadnak szétszórt kristályokat tartalmazó sejtek is. A baktériumsejt a benne lévő mágneses kristálylánc miatt úgy viselkedik, mint egy iránytű, azaz a Föld mágneses tere az erővonalakkal párhuzamos irányba forgatja az egész sejtet. A környező mágneses tér csak a sejt passzív irányulását biztosítja, a baktérium mozgását nem. Valamennyi eddig ismert mágneses baktérium olyan sejtfüggelékekkel (egy vagy több ostorral) rendelkezik, amelyek az aktív mozgást teszik lehetővé.
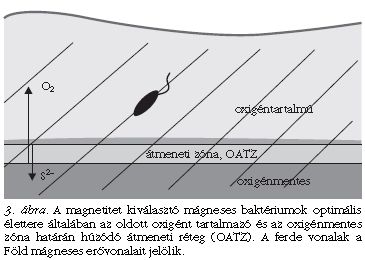
De miért jó egy baktérium számára, ha tájékozódni tud? A választ a baktériumok életterének és fiziológiájának ismeretében adhatjuk meg. A mágneses baktériumok vízi élőhelyeken, tengerekben, tavakban, mocsarakban és ezek üledékeiben élnek, olyan környezetben, ahol az oxigén koncentrációja függőlegesen változik. A mágneses baktériumok többsége számára a túl sok oxigén végzetes, de minimális koncentrációban azért igénylik az oxigént, ezért az oldott oxigént tartalmazó és oxigénmentes rétegek közötti átmeneti zónában (angolul: oxic-anoxic transition zone = OATZ) élnek [3]. Ez a zóna húzódhat egy vízszintes sávban a vízoszlopban (például a Fekete-tengerben ez a helyzet), vagy egy néhány milliméteres rétegre korlátozódhat az üledékben. A legtöbb édesvízi élőhelyen - mint például a Balatonban is - az OATZ a víz-üledék határfelületen vagy kevéssel ez alatt található (3. ábra). Akárcsak mi emberek, akik a földfelszíntől néhány km-es magasságig viseljük el az oxigénhiányt, a mikroaerofil baktériumok pár mm-en belül találják meg a számukra kedvezőéletteret. Az élőlény mérete és az élőhely vastagsága közötti szorzótényező az ember és a baktérium esetében is körülbelül ezerszeres.
A mágneses baktériumok aktív úszásának irányát meghatározza a környező mágneses tér. Ezért a baktériumok kénytelenek a geomágneses erővonalak mentén úszni, amelyek pedig az Egyenlítőtől távolodva egyre nagyobb szögben hajlanak a Föld felszínéhez képest, miáltal a baktériumok mindig "lefelé" vagy "felfelé" úsznak. Ez a viselkedés előnyt jelent az optimális oxigénkoncentráció megtalálása szempontjából, hiszen a sejt egy háromdimenziós keresési problémát egydimenziósra redukál. Nagy oxigénkoncentráció mellett a magnetotaktikus baktériumok az északi félgömbön a mágneses észak felé, míg a déli félgömbön a mágneses dél felé úsznak, azaz mindkét esetben lefelé, a toxikusan nagy oxigénkoncentrációtól távolodva. Az Egyenlítő környékén, ahol a mágneses erővonalak párhuzamosak a felszínnel, közel fele-fele arányban találhatók dél-, illetve északkereső baktérium sejtek [4].
A mágneses baktériumok vizsgálata révén nemcsak magukról a baktériumokról szerezhetünk új ismereteket, hanem az összetettebb élőlények mágneses érzékelésének lehetséges mechanizmusaira is ötleteket gyűjthetünk. A baktériumok sejtjei miniatűr laboratóriumoknak is tekinthetők, amelyekben a mágnességet a nanométeres mérettartományban vizsgálhatjuk. Ezért mind a biológia, mind az ásványtan és az anyagtudomány számára érdekes új eredményeket tartogathat a mágneses baktériumok tanulmányozása.
A mágneses baktériumok irányulásának szükséges feltétele, hogy a bennük lévő kristályláncok állandó mágneses momentummal rendelkezzenek. A kristályok mágneses tulajdonságait több fizikai és kémiai paraméter befolyásolja, amelyek egymással versengő mágneses hatásokat eredményeznek. Ezért a baktériumok sejtjeiben lévő nanokristályok mágnességét csak úgy érthetjük meg, ha részletesen tanulmányozzuk a kristályok összetételét, szerkezetét, alakját és kristálytani irányulását.
A ferrimágneses nanokristályok mágneses tulajdonságai szempontjából a legfontosabb paraméter a kristályméret. A magnetit esetében a 120 nm-nél nagyobb, izometrikus kristályok elméleti számítások szerint mágneses két- vagy többdomének, azaz doménfallal elválasztott, eltérő irányban mágnesezett tartományokat tartalmaznak [5]. A 30-120 nm-es mérettartományban a kristályok egyetlen mágneses domént tartalmaznak. Ezen egydoménes kristályok olyan dipólusok, amelyekben a mágneses indukcióvektor iránya állandó. A 30 nm-nél kisebb kristályok szuperparamágnesesek, azaz a mágneses momentumuk irányát a hőmozgás állandóan változtatja.
A mágnességet a ferrimágneses részecskék alakja is befolyásolja, amely hatást alak-anizotrópiának nevezzük. Minél megnyúltabb egy kristály, annál valószínűbb, hogy a mágneses indukció a hossztengelyével lesz párhuzamos. A magnetokristályos anizotrópia a kristályszerkezet mágnességre gyakorolt hatására utal: vannak a kristályban mágnesesen "lágy" és "kemény" irányok. A magnetit esetén a könnyű mágnesezhetőség iránya a kristálytani [111] irány, azaz a dipólus ebben az irányban hajlamos kialakulni. Az egyes kristályok pozíciója is hat a mágnességükre, mivel a közeli részecskék egymással mágneses kölcsönhatásba lépnek. A mágneses baktériumokban lévő nanokristályokon mindezen tényezők (kristályméret, -alak, kristálytani irányulás, az egyes részecskék elrendeződése) mágnességre gyakorolt hatása látványosan és mennyiségileg tanulmányozható elektronholográfia segítségével.
A transzmissziós elektronmikroszkópos (TEM) elektronholográfia módszerét Gábor Dénes írta le, aki ezért Nobel-díjat is kapott. A módszert a gyakorlatban azonban csak az 1990-es évektől alkalmazzák, mióta megfelelően koherens elektron-sugárforrással felszerelt elektronmikroszkópok is rendelkezésünkre állnak.
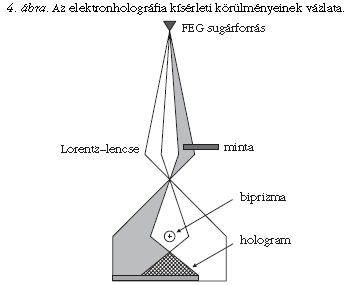
A módszer lényege, hogy a mintán áthaladt elektronsugár fázisát is rögzítjük, nemcsak az intenzitását. A fázis mérése az anyag mágneses tulajdonságainak mennyiségi vizsgálatát teszi lehetővé. Az elektronholográfia kísérleti elrendezését a 4. ábra mutatja: nélkülözhetetlen a koherens elektronsugarat kibocsátó téremissziós katód (angolul: field-emission gun = FEG), a kettősprizma ("biprizma", ami egy Pt- vagy Au-bevonatú huzal, amelyre pozitív feszültség kapcsolható), és a mágneses anyagok vizsgálatához a Lorentz-lencse, amely mágneses tértől mentes leképezést tesz lehetővé. Az elektron-sugárnyaláb egyik része a minta vizsgálni kívánt területén, másik része pedig vákuumon halad át. A mintában a sugár fáziseltolódást szenved a vákuumon áthaladt sugárhoz képest. A kettősprizma sugármenetbe iktatása révén a mikroszkóp ernyőjén a két sugárnyaláb egymással interferál. A keletkező hologram interferenciacsíkjainak periodicitása a fázisra, a csíkok intenzitása az amplitúdóra vonatkozó információt hordozza. Ezen paramétereket a gyakorlatban a hologram Fourier-transzformáltjából lehet megállapítani. A Fourier-transzformált egy maximumhelyének inverz Fourier-transzformálásával kapjuk végül a fázisképet, amelyen a kontraszt a mintát elhagyó elektronsugár fázisának függvénye.
A mintában létrejövő φ(x) fáziseltolódást a közepes belsőpotenciál (angolul: mean inner potential = MIP, ami a helyi összetételtől és sűrűségtől függ), és a B mágneses indukcióvektornak a minta síkjával párhuzamos, a beeső sugár irányában integrált komponense befolyásolja:
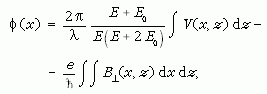
ahol z a beeső elektronsugár iránya, x egy irány a minta síkjában,  a
mágneses indukció x-re és z-re is merőleges komponense, V a minta közepes belsőpotenciálja,
λ az elektronsugár hullámhossza, E és E0 pedig a beeső elektronok mozgási és nyugalmi
tömege [6]. A fenti egyenlet alapján a hologramokon megfigyelhető fáziseltolódás két tagból tevődik össze:
φ(x) = φmip + φmag. A mágnesség tanulmányozásához a φmag,
míg a mintapotenciál, azaz a mintavastagság meghatározásához a φmip ismeretére van szükség.
A két tag fáziseltolódásra gyakorolt hatása egy kísérleti eljárással elkülöníthető.
a
mágneses indukció x-re és z-re is merőleges komponense, V a minta közepes belsőpotenciálja,
λ az elektronsugár hullámhossza, E és E0 pedig a beeső elektronok mozgási és nyugalmi
tömege [6]. A fenti egyenlet alapján a hologramokon megfigyelhető fáziseltolódás két tagból tevődik össze:
φ(x) = φmip + φmag. A mágnesség tanulmányozásához a φmag,
míg a mintapotenciál, azaz a mintavastagság meghatározásához a φmip ismeretére van szükség.
A két tag fáziseltolódásra gyakorolt hatása egy kísérleti eljárással elkülöníthető.
A különválasztott φmag fáziskép alapján mágneses kontúrvonalak számíthatók, amelyek vizuálisan segítik a mágneses tér nagyságának és irányának tanulmányozását (5.e ábra). A mágneses kontúrvonalas ábrák információt adnak a mágneses doménszerkezetről, a kristályok magnetosztatikus, illetve alak-anizotrópiájáról és az egyes részecskék mágneses kölcsönhatásairól. Az elektronholográfia segítségével egyes mágneses tulajdonságok mennyiségileg jellemezhetők. A mágnesség által okozott fáziseltolódásból számítható az egyes nanokristályok és az egész baktériumsejt mágneses momentuma [6].
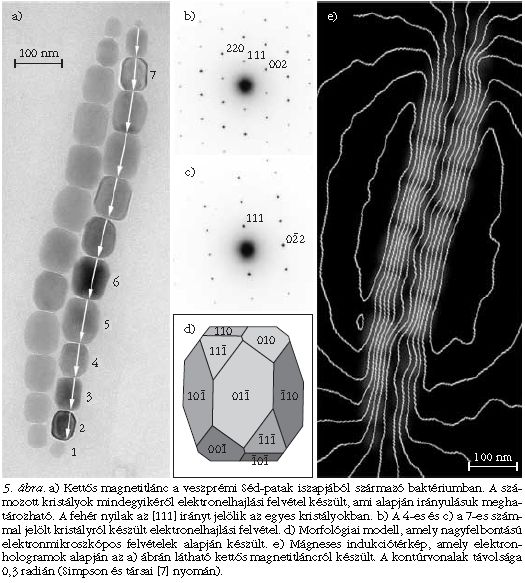
A mágneses baktériumok morfológiailag és a bennük lévő mágneses kristályok jellemzői alapján is sokfélék. Például a Balatonban rendszerint 2-3-féle különböző sejtet lehet találni, ha az üledékből mágnessel dúsítjuk a mintát. Vannak baktériumtörzsek, amelyek kubooktaéderes magnetit-kristályokat választanak ki, míg mások megnyúlt, hasáb morfológiájú kristályokat. A magnetoszómák elrendeződése is sokféle lehet: akadnak egyszeres, kétszeres vagy akár sokszoros kristályláncok, sőt egyes sejttípusok szórt kristályokat tartalmazhatnak (2. ábra). A veszprémi Séd-patak iszapjából olyan coccus morfológiájú baktériumokat gyűjtöttünk, amelyekben a sejt két szemközti oldalán egy-egy kettős magnetitlánc húzódik. Egy ilyen kettős lánc példáján mutatom be az elektronholográfia alkalmazását.
Az 5.a ábra világos látóterű TEM-felvételén látható, hogy a magnetit nanokristályok mind hasonló méretűek és morfológiájúak, kivéve a láncvégi, kisebb kristályokat, amelyek még növekednek. A kristályok 70-100 nm hosszúak, tehát méretük miatt a mágneses egydomén tartományba tartoznak. A magnetit-kristály egyensúlyi morfológiája az oktaéder. Ezzel szemben a képen látható kristályok mind kissé megnyúlt, hasáb alakot mutatnak (5.d ábra), ami rendkívül szokatlan a magnetit esetében, és csakis a kristálynövekedés pontos biológiai szabályozásának köszönhető. A számozott kristályokról orientált elektronelhajlási felvételek készültek (5.b és 5.c ábra), amelyek alapján megállapítható, hogy a kristályok [111] iránya a lánc tengelyével párhuzamos, míg az erre merőleges kristálytani irány tetszőleges. A kristálylánc gyöngyfüzérhez hasonlítható, amelyben rögzített a "zsinórral" párhuzamos irány, de ekörül tetszőlegesen elfordulhatnak a "gyöngyök".
Az 5.e ábrán a kettős magnetitláncról készült mágneses indukciótérkép látható. A térkép több elektronhologram alapján készült. A kontúrvonalak sűrűsége a vonalakra merőleges irányú indukcióváltozással arányos, a vonalak iránya pedig megmutatja az indukció irányát. A képről leolvasható, hogy a kettős lánc egyetlen rúdmágnesként viselkedik. Az egyes nanokristályokban a kontúrvonalak egymással párhuzamosan futnak, tehát minden egyes kristály mágneses egydomén. Valamennyi kristály egy-egy mágneses dipólus, amelyben az indukció a lánc hossztengelyével párhuzamos. A láncvégi apró kristályok a méretüknél fogva szuperparamágnesesek lennének, azonban a térkép szerint ezek is mágnesesek, ami a többi láncbeli kristállyal való mágneses kölcsönhatás következménye.
A mágneses indukciótérkép azt bizonyítja, hogy a baktérium a lehetőleg hatékonyabban "gyártott" iránytűt: a mágneses egydomén mérettartományba tartozó kristályokat szintetizált, amelyek rendezett láncot alkotnak. A kristályok megnyúlási iránya párhuzamos a lánccal, tehát az alak-anizotrópia is a lánccal párhuzamos dipólmomentumot növeli. A könnyű mágnesezhetőségi irány, az [111] minden kristályban párhuzamos a lánccal, azaz a magnetokristályos anizotrópia hatása is hozzáadódik a dipólmomentumhoz. Végül, a kristályok közti mágneses kölcsönhatások is a "tökéletes rúdmágnes" kialakulását segítik elő. A Séd iszapjában élőbaktériumokban tehát a mágnességre ható minden fizikai tényezőhatása összeadódik.
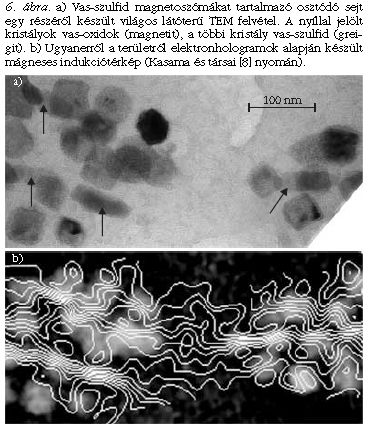
Nem minden mágneses baktérium választ ki a fentihez hasonló tökéletes iránytűt. A vas-szulfid-ásványokat (greigitet) tartalmazó, tengerben élő baktériumokra például jellemző, hogy változatos morfológiájú, némileg rendezetlenül elhelyezkedő kristályokat tartalmaznak (6. ábra). Sőt, egyes esetekben a kristályláncban mind greigit, mind magnetit előfordul. A kétféle ásvány morfológiája eltérő: a 6.a ábrán látható, hogy a nyíllal jelölt magnetit-kristályok erősen megnyúltak, míg a greigit-kristályok izometrikus alakúak. A mágneses indukciótérkép (6.b ábra) jól mutatja, hogy a rendezetlenül elhelyezkedő és tetszőleges kristálytani orientációban lévő greigit-kristályok kanyargós, rendezetlen lefutású mágneses kontúrvonalakat eredményeznek. Az erősebben mágneses és hosszúkás magnetit-kristályokban a kontúrvonalak sűrűbbek és irányuk sokkal jobban definiált (párhuzamos a lánccal), mint a greigit-kristályokban. Ebben a baktériumban tehát nem valósul meg olyan szigorú biológiai szabályozás, mint az 5. ábrán bemutatott magnetitláncnál. Itt a vas-szulfid-kristályok tetszőleges alakja és kristálytani iranya miatt az alak- és a magnetokristályos anizotropia hatása verseng a kristályok közötti mágneses kölcsönhatással.
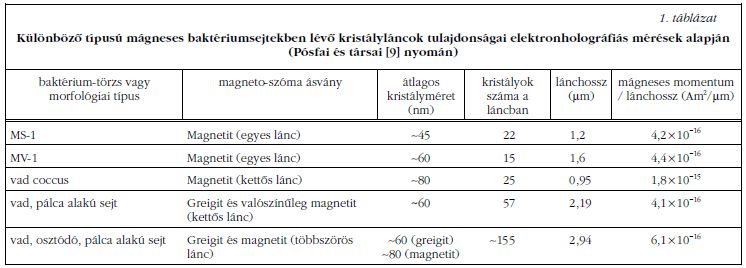
Mind tenyésztett, mind vad típusú, különböző méretű és alakú magnetoszómákat tartalmazó baktériumsejtekben elektronhologramok alapján megmértük a kristályláncok mágneses momentumát. A sejtek mágneses mikroszerkezete közti jelentős különbségek ellenére (amelyeket az 5. és 6. ábra is illusztrál) az egységnyi lánchosszra jutó mágneses momentum minden sejtben közel azonos értéket mutatott (1. táblázat ). Ez arra utal, hogy valamennyi sejt hatékonyan irányul a geomágneses térben. A Langevin-függvény alapján kiszámítható, hogy az ~50 µT erősségű geomágneses térben a vizsgált sejtek az idő ~90%-ában a mágneses erővonalakkal párhuzamosan úsznak. Azon sejtek, melyek rendezetlen irányulású kristályokból álló láncokat választanak ki (6. ábra), a kevésbé hatékony elrendezést azzal egyenlítik ki, hogy sokkal több kristályt növesztenek, mint a tökéletes iránytűt kiválasztó sejtek (5. ábra). A mágneses baktériumok tehát úgy optimalizáltak a biomineralizációs folyamatot, hogy a leghatékonyabban tudjanak a geomágneses térben navigálni.
A mágneses baktériumok sejtjeiben ferrimágneses nanokristályok képződnek. Az egyes sejtek eltérő alakú, méretű és rendezettségű kristályokat választanak ki. A transzmissziós elektronmikroszkópban elektronholografia alkalmazásával a nanokristályok mágnessége mennyiségileg vizsgálható. A különböző mágneses baktériumsejtek elektronholográfiás vizsgálata alapján megállapítható, hogy a nanokristályok fizikai tulajdonságai közül leginkább az alak-anizotrópia befolyásolja a kristályokban a mágneses indukció irányát, míg a magnetokristályos anizotrópia és a kristályok közötti kölcsönhatás kevésbé jelentős. A baktériumokban a nanokristályok egy-egy mágneses domént tartalmaznak, és a sejtek mágneses momentuma elegendően nagy ahhoz, hogy a baktérium a geomágneses tér erővonalaival párhuzamosan irányuljon. Hasonló elektronholográfiás mérések segítségével a jövőben a bonyolultabb élőlényekben (pl. madarakban vagy az emberi agyban) előforduló mágneses kristályok biológiai szerepét is jobban megérthetjük.
Az elektronholográfiás mérések a University of Cambridge, Department of Materials Science and Metallurgy elektronmikroszkóp laboratóriumában készültek Rafal Dunin-Borkowski vezetésével, Takeshi Kasama és Ed Simpson közreműködésével.