
Fizikai Szemle honlap |
Tartalomjegyzék |
Blahó Miklós, Horváth Gábor, ELTE Fizikai Intézet, Biológiai Fizika Tanszék, Budapest
Hegedűs Ramón, Gironai Egyetem, Számítógépes Látás és Robotika Csoport, Girona, Spanyolország
Kriska György, ELTE Biológiai Intézet, Biológiai Szakmódszertani Csoport, Budapest
Gerics Balázs, Szent István Egyetem, Anatómiai és Szövettani Tanszék, Budapest
Farkas Róbert, Szent István Egyetem, Parazitológiai és Állattani Tanszék, Budapest
Susanne Åkesson, Lundi Egyetem, Biológia Tanszék, Lund, Svédország
A fehér lovak gyakran szenvednek az ultraibolya napsugárzással szembeni nagy érzékenységükből fakadó rosszíndulatú bőrrákban és látórendszeri betegségekben. Ráadásul a vadon élő fehér lovakat a ragadozók könnyebben elejtik, mert fehérségük miatt kevésbé tudnak rejtőzködni, mint sötétebb színű fajtársaik. Nagyobb sebezhetőségük ellenére a fehér lovakat az emberek évezredek óta nagy becsben tartják, éppen a természetbeni ritkaságuk miatt. Cikkünkben megmutatjuk, hogy a bögölyök kevésbé vonzódnak a fehér lovakhoz, mint a sötét színűekhez. A bögölyök számos egészségügyi és gazdasági problémát okoznak az embereknek és állatoknak egyaránt, mivel nőstényeik betegségek kórokozóit terjesztik, miközben a gerincesek vérét szívják. Azt is bizonyítjuk, hogy a bögölyök a vérszívásra alkalmas gazdaállatot részben az annak testfelületéről visszavert poláros fény segítségével találják meg. A bögölyök főként fekete és barna szőrű lovakhoz való vonzódása a pozitív polarotaxisukkal, vagyis az erősen és vízszíntesen poláros fényhez való vonzódásukkal magyarázható. Mivel a gazdaállat színe meghatározza a bögölyökre kifejtett vonzerejét is, ezáltal kihat a gazdaállat kórokozók általi megfertőződésére is. Habár kizárólag a bögölyök és lovak közti vizuális kölcsönhatást vizsgáltuk, a jelentős nemzetközi visszhangot kiváltó eredményeink érvényesek lehetnek a polarotaktikus bögölyök más gazdaállataira is.
A természetes kiválasztódás és a hímek versengése a nőstényekért sok esetben vezetett a kültakaró (tollazat, szőrzet) látványos mintázataihoz, amelyek a hímekben hordozott "jó gének" és a paraziták elleni nagy ellenálló képesség vizuális jelzőinek számítanak [1-3]. A tesztoszteron szabályozza a szőrzet növekedését [4] és a melaninképződést [5], valamint befőlyásolja a hímek agresszióját [6], amely jellemzők a hímek dominanciájával és megnövekedett párosodási lehetőségével állnak szőros kapcsolatban. Ugyanakkor a magasabb tesztoszteronszínt nagyobb parazitaterheléssel és a paraziták miatti nagyobb halálozási aránnyal jár együtt [7, 8]. A feltűnőségük miatti nagy sebezhetőségük következtében a természetben csak igen ritkán főrdulnak elő fehér (világos szürke vagy albínó) lovak és más patás állatok. Az ilyen fehér állatok sokkal kevésbé védettek a napsugárzás ultraibolya összetevőjével szemben, ami megnöveli a bőrrák kialakulásának esélyét, továbbá a látórendszer betegségeihez vezethet [9]. Egy fehér patást a ragadozók könnyen észrevesznek, ezért a fehér egyedek az evolúció során kiszelektálódtak a vadonélő populációkból. Másrészt viszont - éppen a természetbeli ritkaságuk miatt - az emberek kitenyésztették a fehér (lovas szaknyelven szürke) lovak különféle fajtáit. A fehér ló az emberi kultúrában idővel a ritkaság, a méltóság és a gazdagság státusz-szimbólumává vált [10]. Gondoljunk csak például Árpád vezérünk vagy Horthy Miklós fehér lovára, illetve a csak a mesékben létező fehér egyszarvúra.
A fehér lovak egy speciális mutációt hordoznak, ami egy több ezer évvel ezelőtt elt közös ősre vezethető vissza [9]. A fehér lovak többsége egy őszülést okozó domináns mutációt hordoz, amennyiben e lovak színes (fekete vagy barna) szörzettel születnek, de egyéves korukra megőszülnek. E főlyamat hasonlít az emberek őszülésére, csak a lovaknál sokkal gyorsabban történik. A ma élő lovak mintegy 10%-a hordozza ezt az őszülést kiváltó mutációt. E genetikai mutáció miatt, az emberi gondoskodás ellenére, a fehér lovak sokkal érzékenyebbek a káros ultraibolya- sugárzással szemben, ami növeli a bőrrák kialakulásának esélyét. A 15 évesnél öregebb fehér (szürke) lovak 75%-ának van valamilyen jóindulatú daganata, ami néhány esetben rosszíndulatúvá alakulhat [9]. Tehát a lovak látványos fehérségének ára az ultraibolya-sugárzás kiváltotta bőrrák nagyobb kockázata.

Cikkünkben megmutatjuk, hogy a fehér lovak a bögölyök számára kevésbé vonzóak, mint a sötétebb színűek. Kísérletekkel és képalkotó polarimetriai vizsgálatokkal támasztjuk alá azt, hogy e jelenség a testfelület fénypolarizáló képességével és a bögölyöknel nemrég fölfedezett pozitív polarotaxissal [11] magyarázható, vagyis azzal, hogy a bögölyök vonzódnak az erősen és vízszintesen poláros fény természetes vagy mesterséges főrrásaihoz. Az, hogy a fehér lovak kevésbé vonzzák a bögölyöket, nagyon előnyös tulajdonság, mert a bögölyök sok szenvedést okoznak a lovaknak, és más emlősöknek is. E rovarok vérszívó nőstényei súlyos betegségek kórokozóinak hordozói [12], főlytonos zaklatásuknak köszönhetően pedig a lovak és más növényevő állatok (például szarvasmarhák) kevesebbet legelnek, miáltal testtömegük és tejtermelésük is jelentősen lecsökken [13]. A vadon élő állatok populációira is negatív hatással van a bögölyök folyamatos támadása miatt megzavart táplálkozás, miáltal a gazdaállatok állóképessége (fitnesze) csökken.

A terepen megfigyeléseket és választásos kísérleteket végeztünk, amelyekben a bögölyök lovakhoz, lómakettekhez es különböző fénypolarizáló-képességű tesztfelületekhez való vonzódását tanulmányoztuk. A kapott eredmények szerint nem önmagában a szőrzet sötét színe, hanem fénypolarizáló-képessége okozza a bögölyök erősebb vonzódását a sötétebb lovakhoz. Hogy a választásos terepkísérleteinkben használt lómakettek es tesztfelületek polarizációs sajátságait összehasonlíthassuk a bögölyök által támadott tipikus gazdaállatokéival, élő lovak és szarvasmarhák polarizációs mintázatait is mertük képalkotó polarimetriával. A lovasok, valamint a lovak és más haszonállatok tartói számára is fontos eredményeinket [14] számos más nyomtatott és elektronikus médium mellett a Nature [15], a Science [16] és a Discover Magazine [17] is méltatta.
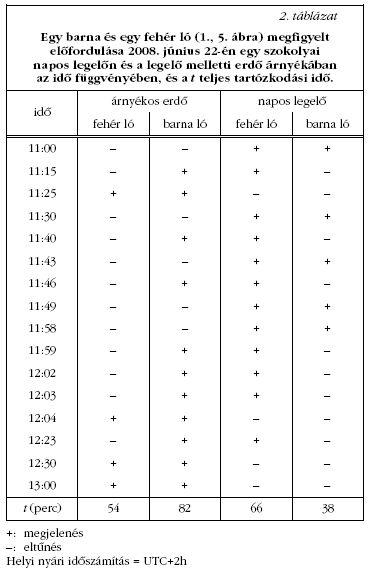
Lovak bögölyök általi zaklatásának megfigyelése
Egy meleg napsütéses napon, 2008. június 22-én
Szokolyán (47° 52' Ny, 19° 00' K) egy legelőn figyeltük
meg egy barna (lovas szaknyelven pej) és egy
fehér (szürke) ló bögölytámadásokkal szembeni viselkedését.
A bögölyök mindkét lovat folyamatosan zaklatták,
a lovakat valosagos bögölyfelhő vette körül.
Hogy a bögölyfelhő méretet valamiképpen dokumentáljuk,
70-70 fényképet készítettünk a legelő barna és
fehér lóról (1. ábra). A 10 megapixeles felbontású
képeken egy számítógépes képernyőn leszámoltuk a
vizuálisan fölismert bögölyöket (1. táblázat ). Az intenzív
bögölytámadások elől a lovak időnként a napos
legelőről a rétet határoló erdő árnyékába menekültek.
Mertük mindkét ló esetén a legelőn és árnyékban
töltött időt (2. táblázat).
 |
 |
Az 1. kísérlet
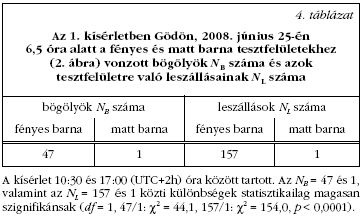
2008. június 23-24-én, illetve 25-26-án zajlott meleg,
napos időben egy-egy lovastanyán Szokolyán
(47° 52' Ny, 19° 00' K) és Gödön (47° 43' Ny, 19° 09'
K), naponta 9:30-tól 17:30-ig (helyi nyári idő =
UTC+2h). Egy adott helyszínen az első napon két
azonos méretű (150 × 150 cm) és minőségű, száraz,
matt, barna vásznat fektettünk vízszintes deszkákra,
egymástól 1 m távolságra. Az egyik vásznat egy színtelen,
átlátszó, 3 mm vastag műanyag lappal fedtük le
(2. ábra). A tesztfelületeket óránként fölcseréltük,
továbbá kétóránként az árnyékból a közeli napos
területre helyeztük át. A tesztfelületek hőmérsékletet
digitális kontakthőmérővel mértük. A két tesztfelület
hőmérséklete a kísérlet során mindvégig - ±0,25°C
pontossággal - azonos volt. Mindkét tesztfelületnél
számoltuk az odavonzott bögölyöket és a tesztfelületre
való leszállásaikat (3. és 4. táblázat ). A második
napon is elvégeztük ezt a kísérletet azzal a különbséggel,
hogy a műanyag lappal borított barna vásznat
egy azonos méretű matt fehér vászonra cseréltük.
A 2. kísérlet
Először 2008. augusztus 3-án 9:00 órától 19:00
óráig (UTC+2h) végeztük el egy lovasiskola melletti
temetőben, Kiskunhalason (46° 43' Ny, 19° 05' K). A
kísérleti elrendezés az 1. kísérlettel megegyező volt,
csak a vízszintes átlátszó, színtelen műanyag lapot
színtelen, szagtalan ragasztóval (Bábolna-Bio® egérragacs)
vontuk be, amely a felületre szálló minden rovart
megfogott, továbbá a tesztfelületek végig árnyékban
voltak. Számoltuk a matt barna felületre szálló és
a csillogó ragadós felülettel csapdába ejtett bögölyöket.
A ragasztóba ragadt bögölyöket azonnal eltávolítottuk
a felületről, így a következő bögöly számára a
csapdába ejtett fajtárs látványa nem befolyásolta a
felület vonzóképességet. A kísérletet 2008. augusztus
4-en ugyanott megismételtük azzal a különbséggel,
hogy a műanyag lappal borított matt barna vásznat
egy azonos méretű matt fehér vászonra cseréltük.
Megint számoltuk a két tesztfelületre szálló bögölyöket.
A tesztfelületek hőmérséklete a kísérlet során
végig azonos volt, mivel árnyékban voltak.

A kísérlet első részét, amiben matt barna és fényes barna tesztfelületeket használtunk, 2009. augusztus 16. és 25. között, naponta 12:00-tól 13:00-ig, összesen 10 alkalommal megismételtük. Számoltuk a vízszintes matt barna felületre szálló és a vízszintes csillogó ragadós felülettel csapdaba ejtett bögölyöket (5. táblázat). A fényes barna felületre ragadt bögölyöket most is azonnal eltávolítottuk. Miközben a ragadós felületről leszedtük a bögölyöket, azok súlyosan megsérültek, ami a faji meghatározásukat lehetetlenné tette. Mindazonáltal bizonyosan bögölyök (Tabanidae: Diptera) voltak. 2009. augusztus 16. és 25. között, naponta 13:00-tól 14:00-ig a kísérlet második felet is - amiben egy száraz matt barna és egy száraz matt fehér tesztfelületet használtunk - tízszer megismételtük, számolva a tesztfelületekre rászálló bögölyöket.
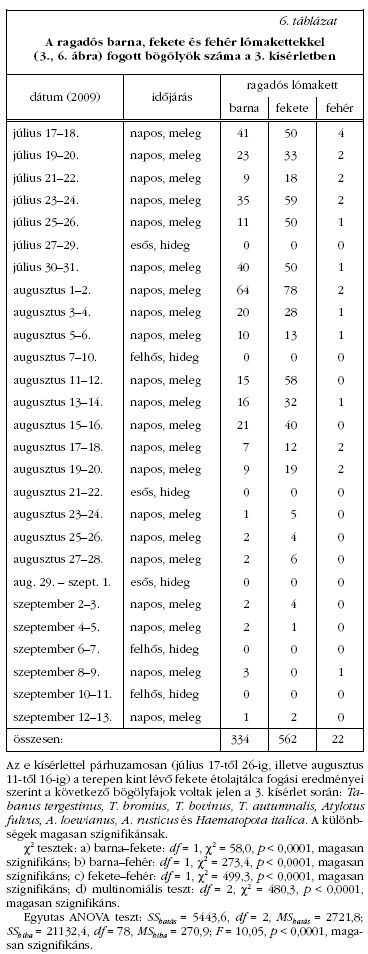
A 3. kísérlet
2009. július 17-től szeptember 13-ig folyamatosan
zajlott egy lovastanyán, Szokolyán (47° 52' Ny, 19°
00' K). Egy barna, egy fekete és egy fehér lómakettet
(mindhárom azonos formájú és méretű: hosszúság:
160 cm, magasság: 110 cm, szélesség: 60 cm) állítottunk
a füves talajra normál testtartásban (3. ábra). A
három lómakettet 5 m távolságra helyeztük el egymástól
egy egyenes mentén. A felületüket minden
második napon színtelen, szagtalan ragasztóval (Bábolna-Bio®
egérragacs) kentük be. A lómakettek sorrendjét
kétnaponként véletlenszerűen fölcseréltük. A
kísérlet helyét úgy választottuk meg, hogy mindhárom
lómakett egyszerre legyen napon vagy árnyékban.
A lómakettek ragadós felületeivel csapdába ejtett
bögölyöket kétnaponta leszedtük és megszámoltuk
(6. táblázat ). Felhős vagy esős időben a bögölyök
nem repültek, s ekkor a lómakettek egyetlen bögölyt
sem fogtak. Miközben a lómakettek ragadós felületéről
a bögölyöket leszedtük, azok súlyosan megsérültek,
ami a faji meghatározásukat lehetetlenné tette.
Mindazonáltal a leszedett rovarok bizonyosan bögölyök
(Tabanidae: Diptera) voltak.

A 4. kísérlet
egy gödi lovastanyán (47° 43' Ny, 19° 09' K) zajlott
2008. július 11. és szeptember 7. között. Öt (fehér, világosszürke,
középszürke, sötétszürke, fekete) étolajjal
töltött tálcát tettünk a földre (4. ábra), sorrendjüket
naponta, véletlenszerűen
változtatva. Éjszakára, valamint
esős időben a tálcákat
letakartuk. A tálcák által csapdába
ejtett bögölyöket begyújtottuk
és etil-alkoholban
tartósítottuk a későbbi határozás
céljából.
Polarizációs mintázatok
mérése
A tesztfelületek (2., 4. ábra),
lómakettek (6. ábra), valamint
lovak és szarvasmarhák
(7. ábra) polarizációs
mintázatait képalkotó polarimetriával
[18] mértük a spektrum
vörös (650 ± 40 nm = a
polarimeter CCD-detektora
maximális érzékenységének
hullámhossza ± annak félérték-szélessége),
zöld (550 ± 40
nm) és kék (450 ± 40 nm) tartományában.

A továbbiakban csak a kék tartományban mért polarizációs mintázatokat mutatjuk be, a mintázatok mindhárom csatornában gyakorlatilag azonosak voltak. A polarizációs méréseket tiszta égbolt alatt végeztük. A polarotaktikus vizirovarok, például a bögölyök akkor érzékelnek víznek egy felületet, ha az arról visszavert fény (i) p linearis polarizációfoka nagyobb egy fajfüggő p* küszöbértéknél, és (ii) az α polarizációirányának a vízszintestől (α = 90°) való Δα = |90°-α| eltérése pedig kisebb egy fajfüggő Δα* küszöbértéknél. Példaként a 6. ábrán p* = 10% és Δα* = 10° értékeket használtunk. Habár e küszöbértékeket önkényesen választottuk, más értékek alkalmazása sem befolyásolta az eredményeket és következtetéseket.
A bögölyök meghatározása
Az etil-alkoholban tartósított
bögölyök faji meghatározását
az egyik szerző (F. R.)
végezte. A 3. kísérlettel párhuzamosan
egy másik kísérletet
is végeztünk július 17-től
26-ig, illetve augusztus 11-től
16-ig. Egy fekete, négyzet
alakú (50 × 50 cm), étolajjal
töltött műanyag tálcával fogtunk
bögölyöket, ami lehetővé
tette az adott területen
jelenlévő bögölyfajok meghatározását
(6. táblázat).

Megfigyeltük, hogy a barna és fehér lovakat a bögölyök egyaránt és folyamatosan támadták (1., 5. ábra). A lovak jellemző védekező reakciói a bögölyökkel szemben a következők voltak: legyezés (farokkal történő ide-oda csapkodás), dobbantás, hempergés a földön, hirtelen megrázkódás, fej himbálása, a vérszívó bögölyök harapással és nyalással történő eltávolítása a kültakaróról (5.d-i ábrák). E viselkedési elemekkel próbálták elhajtani a lovak a rájuk szálló bögölyöket. 70 képpáron számláltuk meg a vizuálisan fölismert bögölyöket a barna és a fehér lovakon, illetve azok körül (1. ábra, 1. táblázat ). A barna lovon 405/110 = 3,7-szer annyi bögölyt számoltunk, mint a fehéren, amely különbség statisztikailag magasan szignifikáns. A barna lónak a fehérhez képesti, bögölyökre kifejtett nagyobb vonzereje összhangban áll a 3. kísérlet eredményével (lásd később).
A terepen azt is megfigyeltük, hogy a nyílt legelőn és az árnyékos erdőben nem azonos a bögölytámadások intenzitása, miáltal a lovak a két terület között folyamatosan ingáztak (2. táblázat). Bizonyos, legeléssel töltött idő után a lovak az árnyékos erdőbe menekültek a bögölyök elől, ahová a bögölyök csak ritkán követték őket, így nyugodtan tudtak pihenni. Egy idő elteltével a lovak újra előmerészkedtek a napos mezőre legelni, ahonnan azonban hamarosan megint visszakényszerültek az árnyékba. Ezt az ingázó viselkedést ismételgették periodikusan a nap közepéig (13:00), mikortól a bögölyök általi zaklatás oly mértékű lett, hogy a lovak már egyáltalán nem tudtak a mezőn legelni. A napos legelőről elsőként mindig a barna ló menekült az erdőbe (2. táblázat ). A barna ló 82 perc, illetve 38 perc, azaz 2,2-szer több időt töltött a bögölymentes, árnyékos erdőben, mint a napos legelőn, míg a fehér ló 65 perc / 54 perc = 1,2-szer több időt töltött a legelőn, mint az árnyékban (2. táblázat).
Mivel nemrég bebizonyosodott, hogy számos bögölyfaj vonzódik az erősen és vízszintesen poláros fényhez [11], föltételezhető volt, hogy a sötétebb (barna) ló nagyobb vonzóképessége részben magyarázható az állat szőrzetének fénypolarizáló-képességével, ami erősen függ annak színétől és világosságától, ahogyan az a 6. és 7. ábrákon is látható. E föltevést négy különböző választásos kísérlettel ellenőriztük, amelyek közül néhányat többször is megismételtünk.

Az 1. kísérletből kiderült, hogy a matt barna vászon, amely a lovak barna szőrét utánozta, a bögölyök számára nem volt vonzó, míg az átlátszó, színtelen műanyaglappal letakart matt barna vászon számos bögölyt vonzott (Szokolyán 44 bögölyt 174 leszállással, Gödön 47 bögölyt 157 leszállással a 3. és 4. táblázatok szerint). E különbségek a megvilágítási körülményektől (napos vagy árnyékos) függetlenül statisztikailag magasan szignifikánsak. A bögölyök azonnal vagy 2-15 felületérintés után szálltak rá az adott felületre. A bögölyök a barna tesztfelületeknél ugyanazt a viselkedést mutatták, mint a bögölyök pozitív polarotaxisának fölfedezésekor korábban lefolytatott választásos kísérletekben [11]. Mikor az 1. kísérletet matt barna és matt fehér felületekkel ismeteltük meg, azok egyáltalán nem vonzottak magukhoz bögölyöket.
Hogy az 1. kísérletben elkerülhetetlenül föllépő pszeudoreplikációt kizárjuk (azaz, hogy ugyanazt a bögölyt többször is megszámoljuk annak többszöri visszatérésekor), elvégeztük a 2. kísérletet, amiben a bögölyök többségét egy ragasztó megfogta. A 2. kísérlet eredménye tovább erősítette az 1. kísérletet. 2008-ban az átlátszó, színtelen, ragacsos műanyag lappal letakart barna felület 21 bögölyt fogott, míg a matt barna és matt fehér felületekre csak egyetlen bögöly szállt le (Χ2 = 18,2, df = 1, p < 0,0001). E kísérletet 2009-ben tízszer megismételve, a következő eredmény adódott: az 5. táblázat szerint ekkor a fényes, ragacsos barna felület 189 bögölyt fogott, míg a matt, száraz barna mindössze négyet. Mindkét különbség statisztikailag magasan szignifikáns. Mikor 2009-ben a 2. kísérletet is 10-szer elvégeztük a száraz matt barna és száraz matt fehér tesztfelületekkel, megint egyikük sem vonzott egyetlen bögölyt sem.
Az 1. és 2. kísérletekből az a következtetés vonható le, hogy a bögölyök nem vonzódnak a vízszintes matt barna (kevésbé poláros) és a matt fehér (depolarizáló) tesztfelületekhez, viszont erősen vonzódnak a vízszintes fényes barna (vízszintesen poláros fényt visszaverő) felületekhez. Tehát a barna lovak bögölyökre kifejtett nagyobb vonzóképessége nem magyarázható pusztán a lovak szőrének színével és fényességével.
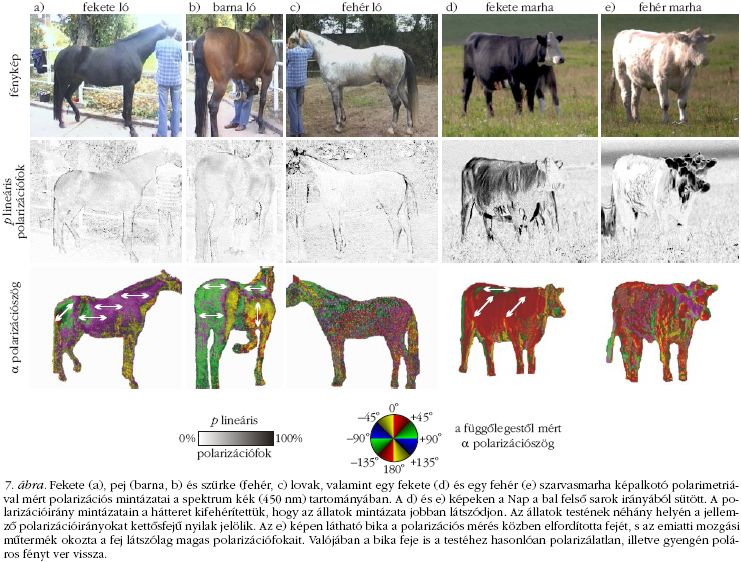
A képalkotó polarimetriai mérések szerint, míg a vízszintes fényes (száraz vagy ragadós) barna tesztfelületek (2. ábra), a szoláris meridiánhoz képesti nézőiránytól függetlenül, mindig erősen és vízszintesen poláros fényt vernek vissza, addig a vízszintes matt száraz barna és fehér felületekről mindig csak gyengén poláros fény verődik vissza, aminek polarizációiránya függ a Naphoz képesti nézőiránytól. Így a felületek bögölyökre kifejtett vonzását nem okozhatja önmagában a barna szín, a visszavert fény polarizációs tulajdonságai is számítanak. E következtetést a 3. kísérlet (3., 6. ábra, 6. táblázat ) eredményei is alátámasztották. A 6. táblázat szerint a fényes ragadós barna és fekete lómakettek 334/22 = 15,2-szer, illetve 562/22 = 25,5-ször annyi bögölyt fogtak, mint a ragadós fényes fehér lómakett. E különbségek most is statisztikailag magasan szignifikánsak. A megfogott bögölyök eloszlása a lómakettek testfelületen véletlenszerű volt, egyik részük sem volt kitüntetett a bögölyök számára. A 6. ábrán látható, hogy a barna és a fekete lómakett egyes részei nagyon erősen poláros fényt tükröznek, míg a fehér lómakett gyakorlatilag polarizálatlan fényt ver vissza. A barna és a fekete lómakett háta, valamint fara ver vissza erősen és vízszintesen poláros fényt (6. ábra 3. sora), igy e testrészek lehetnek a bögölyök számára nagyon vonzóak.

A következő bögölyfajok estek csapdába: Atylotus loewianus, Haematopota italica, Tabanus bovinus, T. maculicornis, T. tergestinus. Az Nfekete - Nsötétszürke (Χ2 = 35,44, df = 1, p < 0,0001) és az Nsötétszürke - Nközépszürke (Χ2 = 14,44, df = 1, p < 0,0001) közti különbségek statisztikailag magasan szignifikánsak, amit * jelöl, míg a fehér, világosszürke és középszürke közti különbségek nem szignifikánsak: Nfehér-Nvilágosszürke (Χ2 = 0,2, df = 1, p = 0,6547), Nfehér-Nközépszürke (Χ2 = 1, df = 1, p = 0,3171). |
A 4. kísérlet (4. ábra) eredményei szerint minél sötétebb egy színtelen, vízszintesen polarizáló tesztfelület, annál vonzóbb a bögölyök számára: a fehér , világosszürke és középszürke olajtálcák a teljes fogásnak csak 0,9-2,7%-át adták, a sötétszürke tálca a bögölyök 19,8%-at fogta meg, míg a fekete talca 74,8%-ot (7. táblázat). Ez az eredmény azzal magyarázható, hogy minél sötétebb egy színtelen (fekete, szürke, fehér ) felület, annál nagyobb a róla visszavert fény polarizációfőka a hullámhossztól függetlenül (4. ábra).
A választásos terepkísérleteinkből és polarizációs méréseinkből az a következtetés vonható le, hogy a nemfémes (szigetelő) felületek közül - függetlenül attól, hogy naposak vagy árnyékosak, barnák vagy színtelenek - a bögölyök számára az a legvonzóbb, amelynek a felületéről visszavert fény vízszintesen poláros és polarizációfőka a legnagyobb. Ennek alapján logikus azt föltételezni, hogy a sötét színű lovak vízszintesen polarizáló testfelületei a bögölyök számára sokkal vonzóbbak, mint a világos színű lovak ugyanazon testrészei. A 4., 6. és 7. ábrák szerint minél sötétebb a testfelület, annál nagyobb a róla visszavert fény polarizációfőka. A fekete, barna, illetve fehér testfelszín erősen, közepesen, illetve gyengén poláros vagy polarizálatlan fényt ver vissza. A nézőiránytól és a megvilágítási körülményektől függően a normál, álló testtartású lovak nyaka, háta és fara általában vízszintesen poláros fényt ver vissza, míg a test többi részéről ferdén vagy függőlegesen poláros fény verődik vissza (a 7. ábra polarizációirány-mintázatain kettősfejű nyilakkal jelölve). E polarizációs sajátságok általánosak és a bögölyök minden gazdaállatára egyaránt érvényesek. Azon fő eredményünket, hogy a polarotaktikus bögölyök kevésbé vonzódnak a fehér lovakhoz (és más világos szőrű emlőshöz) és nagyon vonzódnak a fekete lovakhoz (és más sötét szőrű emlőshöz), a 8. ábra szemlélteti.
Napsütésben a sötétebb lovak szőre jobban fölmelegszik, mint a világosabbaké, mert a sötétebb testfelület több fényt nyel el. Ez részben magyarázhatna, hogy a sötétebb lovak miért vonzanak több bögölyt, ha a bögölyök egyértelműen vonzódnának a melegebb helyekhez. Habar a bögölyöknél a gazdaállat kiválasztásában a testhőmérséklet is szerepet játszik [19], a gazdaállat testfelületének hőmérsékletét csak akkor tudjak érzékelni, ha mar rászálltak, mert nagyobb távolságból a különböző színű gazdaállatok testfelszíni hőmérséklete a repülő bögölyök számára észlelhetetlen, mivel nincsen infravörös látásuk. Továbbá, az 1. és 2. kísérletünkben a különböző tesztfelületek hőmérséklete mindig azonos volt. Ezek alapján állíthatjuk, hogy a sötét szőrű lovak bögölyökre kifejtett nagyobb vonzóképessége nem magyarázható csupán a sötét és világos lovak testfelszíni hőmérsékletének esetleges különbségével.
Habár egy barna lónak lehetne a bögölyök számára vonzóbb szaga (például a sötétebb testfelszín magasabb hőmérséklete miatti erősebb izzadástól), a választásos kísérleteinkben használt tesztfelületeknek egyforma volt a szaga. Ezért a különböző színű lovak szagának esetleges különbsége nem lehet a bögölyök preferenciájának fő oka. Mivel a vizsgált pej (barna) és szürke (fehér ) lovak alakjában és mozgásában sem volt fölfedezhető semmilyen lényeges különbség, ez sem magyarázhatja, hogy a bögölyök miért vonzódnak jobban a barna lovakhoz. Az 1. és 2. kísérletünk együttes eredményei kizárják annak lehetőségét is, hogy a barna szín önmagában jobban vonzaná a bögölyöket.

A föntiek alapján az egyetlen lehetséges magyarázat arra, hogy a barna lovak több bögölyt vonzanak a fehéreknél, az a testfelületükről visszavert fény polarizációja. Az 1. és 2. kísérletből kiderült, hogy a bögölyök jobban vonzódnak az erősen és vízszintesen poláros fényt visszaverő fényes barna tesztfelülethez, mint a gyengén és nem mindig vízszintesen poláros fényt visszaverő matt barna vagy a depolarizáló matt fehér tesztfelületekhez. Ezért nem a szín és/vagy a fényesség a döntő, hanem a célpont (tesztfelület, gazdaállat) polarizációs mintázata.
Az evolució során kifejlődött a bögölyök polarizációlátása és pozitív polarotaxisa, amely képesség a víz felszínéről visszavert fény vízszintes polarizációjának köszönhetően segíti őket a vízkeresésben. A víz közvetlen környezete ideális helye a bögölyök petézésének, a fajtársakkal való találkozásnak, valamint az inni és/vagy fürdeni rendszeresen a vízhez látogató gazdaállatok megtalálásának is. Logikus föltételezni, hogy a bögölyök polarotaxisa a gazdaállat kiválasztásában is szerepet játszik. Ha egy bögöly választhat egy sötét és egy világos megjelenésű, s minden egyéb tulajdonságában (nem, alak, méret, szag, hőmérséklet, mozgás, ver stb.) megegyező gazdaállat közül, akkor a sötétebbet választja, mert a sötétebb gazdaállatról visszavert fény polárosabb. Ezek alapján a következő föltevések tehetők:
A 4. kísérletünk eredményei szerint a vízszintesen poláros fényt visszaverő színtelen felületek közül a fényt jobban polarizálók több bögölyt vonzanak, mint a gyöngébben polarizálók. Ez megerősíti a 2. föltevést. A képalkotó polarimetriás mérések (6., 7. ábra) megmutattak, hogy a sötétebb szőrű lovak és szarvasmarhák nagyobb polarizációfokú fényt vernek vissza, és ezen álló állatok nyaka, háta és fara vízszintesen poláros fényt verhet vissza. Habár a gazdaállatról visszavert fény polarizációiránya függ az állat testtartásától, a megvilágítási viszonyoktól és a szoláris meridiánhoz képesti nézőiránytól, a napfénnyel megvilágított vízszintes állati testfelület mindig vízszintesen poláros fényt ver vissza, ha a szoláris vagy az antiszoláris meridián irányából nézzük. Mindezek alátámasztják a 3. föltevést. Az 1. föltevés pedig a következőkön alapszik: a vizsgált bögölyöknek pozitív polarotaxisa van [11]. Az 1. föltevés teljesüléséhez például az kell, hogy a 2. és 3. föltevések összhangban legyenek a megfigyeléseink és kísérleteink eredményeivel. A vadonban a bögölyök gazdaállatainak, a nagytestű növényevőknek, általában sötét (barna, szürke, fekete) a szőrzete. Álló helyzetben ezen állatok nyaka, háta és fara mindig mérsékelten (barna, szürke) vagy erősen (fekete) és vízszintesen poláros fényt ver vissza. Így tehát a bögölyök a pozitív polarotaxisukkal e gazdaállatokat éppúgy meg tudják találni, mint a vízfelületeket. Mai tudásunk szerint nem létezik más magyarázata annak a jelenségnek, hogy a bögölyök miért vonzódnak jobban a sötétebb színű gazdaállatokhoz, mint a világosakhoz.
Mivel a bögölyök számára a sötét szőrű állatok világos szőrűekkel szembeni előnyben részesítése semmilyen előnyt vagy hátrányt nem jelent a természetes környezetben, e preferencia a bögölyök pozitív polarotaxisának egy semleges következményeként tekinthető. Másrészt viszont a gazdaállatok szemszögéből nézve, a bögölyök szóban forgó sötétpreferenciája nem semleges: a sötét szőrű állatoknak hátrányos, a világos szőrűeknek pedig előnyös a kisebb fertőzésveszélynek és kevesebb zaklatásnak köszönhetően. Az ember által mesterségesen tenyésztett és tartott lovak esetében a legkevésbé bögölyvonzó fehér testfelszín előnye részben ellensúlyozhatja az ultraibolya-sugárzás okozta bőrrákra való nagyobb érzékenységet. A vadon élő lovaknál mindazonáltal a sötétebb testfelület (a könnyebb rejtőzködés miatt) előnyösebb, mint a világos (amely a bögölyök számára kevésbé vonzó).
Kísérleti eredményeink határozottan alátámasztják azt, hogy a gazdaállatok testfelületének fehérsége egy bizonyos szempontból előnyös tulajdonság, mivel a fehér állatok a bögölyöket kevésbé vonzzak, mint a sötétek. Szamos bögölyfaj pozitív polarotaxissal bír, azaz vonzódik az erősen és vízszintesen poláros fényhez [11, 20], csakúgy, mint a vizirovarok általában [18, 21-23]. A bögölyök pozitív polarotaxisának a következő biológiai szerepei lehetnek:
A föntiek alapján arra a következtetésre juthatunk, hogy a bögölyök polarotaxisának a gazdaállat kiválasztásában is fontos szerepe van. Habar a különböző színű gazdaállatok egyaránt megfelelőek a nőstény bögölyöknek a vérszíváshoz, a bögölyök mégis a sötétebb (barna, fekete) szőrű állatokat részesítik előnyben a világos (szürke, fehér ) szőrűekkel szemben. Ez a jelenség jól magyarázható a gazdaállatok testfelületének fénypolarizáló-képességével és a bögölyök polarotaktikus viselkedésével. A korábbi föltevések szerint a bögölyök gazdaállat-kiválasztásában csak a mozgás, alak, szín, fényesség, szag és hőmérséklet játszik fontos szerepet [19, 24-26].
A 3. kísérletben a bögölyök a lómakettekről (3., 6. ábra) visszavert vízszintesen poláros fény mennyiségének megfelelően vonzódtak azokhoz. Habár nagy távolságból nézve a vízszintesen poláros fényt visszaverő testrészek lehetnek a polarotaktikus bögölyök számára a legvonzóbbak, a bögölyök eloszlása a lómakettek teljes felületen véletlenszerű (egyenletes) volt. Ez azt mutatja, hogy a vízszintesen poláros fény mellett más vizuális jeleknek (a gazdaállat színének, fényességének és testformájának) is fontos szerepe van a gazdaállat megtalálásában és kiválasztásában. Habár csak a bögölyök és a lovak vizuális kölcsönhatását vizsgáltuk, a kapott eredmények más gazdaállatokra is érvényesek lehetnek, mivel a szóban forgó vonzás alapja, a pozitív polarotaxis, általánosnak tűnik a bögölyöknél [11], továbbá az azonos világosságú, színű és felületi érdességű, de különböző fajú gazdaállatok polarizációs mintázatai gyakorlatilag egyformák.
A sötét színű, kényes lovakat tartó gazdáknak végezetül azt tudjuk javasolni, hogy azon fekete vagy barna lovakat, melyeket nagyon zaklatják a vérszívó bögölyök, takarják le valahogyan egy vékony, matt fehér vászonnal, ami csak gyengén poláros vagy polarizálatlan fényt ver vissza, miáltal kevésbé vonzza a bögölyöket.
Kutatásainkat az Európai Unió (Eu-FP7, TabaNOid-232366: Trap for the novel control of horse-flies on open-air fields) és az OTKA (K-6846: Közvetlen és közvetett polarotaxis vizsgálata tegzeseknél és kétszárnyúaknál) is támogatja.
