Fizikai Szemle honlap |
Tartalomjegyzék |
Kertész Krisztián, Piszter Gábor, Vértesy Zofia, Biró László Péter
MTA TTK Műszaki Fizikai és Anyagtudományi Intézet1
Bálint Zsolt
Magyar Természettudományi Múzeum2
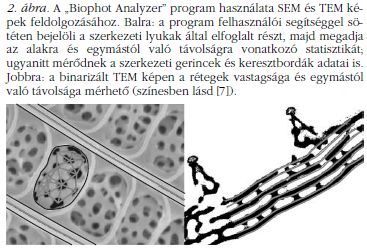
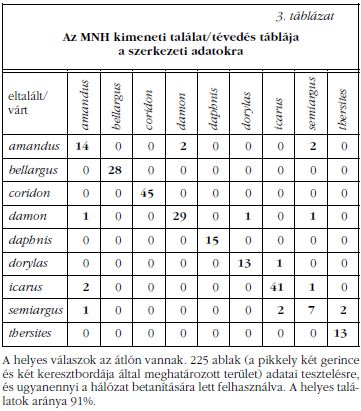
Amint az a SEM és TEM felvételeken látható ( 1. ábra az előző szám hátsó belső borítóján), minden vizsgált lepkefaj pikkelyeiben jellegzetes szivacsos szerkezet található. A képeket szemlélve megfigyelhetünk eltéréseket, de pontos mennyiségi különbségek megállapítása lehetetlennek látszik. A SEM képekből a pikkelygerincek, keresztbordák és a legfelső lyukacsos réteg méretei nyerhetők ki, míg a mélységi adatokhoz a TEM képeket használhatjuk. A szivacsszerű nanoszerkezet adatai az "ablak"-nak nevezett egységből (két gerinc és két keresztborda által meghatározott terület) származnak. Az összes adat három csoportra osztható: gerincek és keresztbordák távolságai és szélességei, a szivacsos szerkezet jellemzői, valamint a TEM képekről leolvasható rétegvastagságok és távolságok (2. ábra). Így összesen 24 adat nyerhető, de a további vizsgálataink szerint 9 paraméter (2. táblázat) elégséges az MNH (mesterséges neurális hálózat) elemzéshez. Összegezve, a 9 faj pikkelyein 450 ablakot mértünk meg, ezek fele az MNH-tanításban vett részt, a többi a tesztelésben. Az eredmények szerint (3. táblázat) a neurális hálózat a 255 ablakból 205-öt helyesen határozott meg, ami 91%-os találatot jelent.
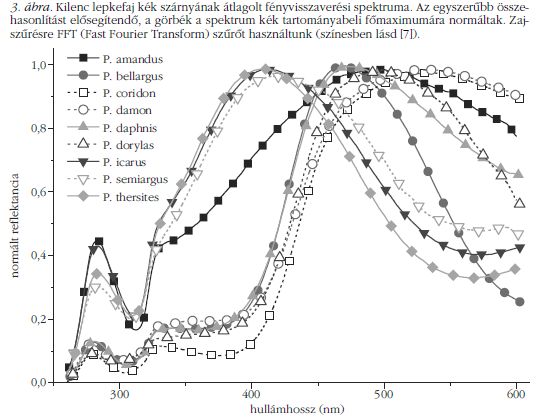
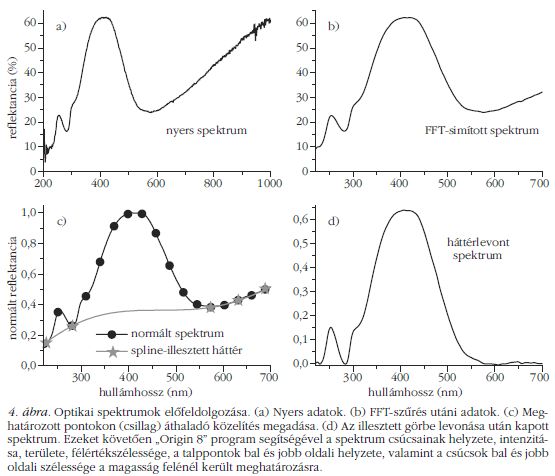
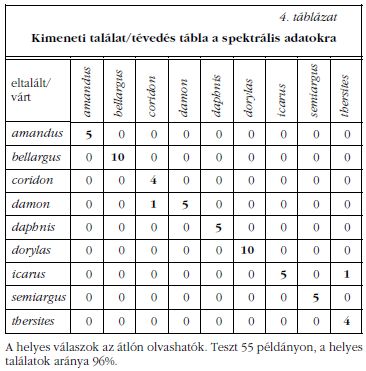
Korábban megmutattuk, hogy egy adott fajhoz tartozó egyedek színe gyakorlatilag megegyezik [8]. A fajok spektrális megkülönböztetésének automatizálását a normált és átlagolt visszaverési spektrumokon (3. ábra) végeztük. A spektrumok jellemzőit (4. ábra) a következő paraméterek határozzák meg: a csúcs maximumának hullámhossza, az intenzitás ebben a pontban, a csúcs területe és félértékszélessége, a csúcs talppontjainak helyzete és a félértékszélesség két oldalának értéke. Ez 8 paraméter csúcsonként, ami összesen a három csúcsra (a főmaximum, valamint balra két kisebb intenzitású csúcs) 24 bemeneti adatot jelent a mesterséges neurális hálózatba. Minden fajból legkevesebb 10 példányt mértünk, amelynek fele vett részt az MNH tanításában. Az eredményeket a szerkezeti vizsgálathoz hasonlóan közöljük (4. táblázat), a spektrumok alapján végzett faj szerinti osztályozásban a találati pontosság 96% volt.

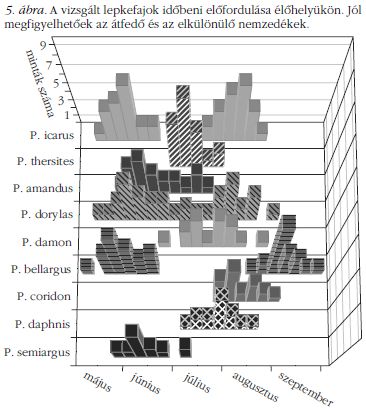
A rajzási időszakok meghatározására a lepkék befogási idejét használtuk fel. Ehhez a Természettudományi Múzeum Állattárában feljegyeztük 285 lepke befogási idejét (hónap, nap), és ezt egy hisztogramban ábrázoltuk (5. ábra). A lejegyzett minták nagy száma szükséges az eloszlás követése és az évenkénti eltérő éghajlati viszonyok hatásának átlagolása miatt.
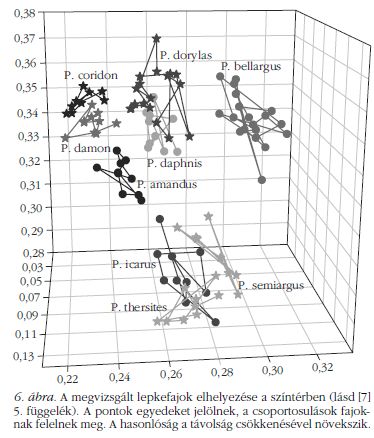
A színek megkülönböztetésére használt egyik ábrázolási mód a CIE (Commission internationale de l'éclairage) színinger-háromszög [10], amely a színek emberi látáshoz illeszkedő kétdimenziós megjelenítése. Korábban már végeztünk ehhez hasonló vizsgálatot [8], viszont figyelembe kell venni, hogy a vizsgált lepkék nem az emberre jellemző háromféle szín érzékelésére képes fotoreceptorokkal rendelkeznek, hanem ez kiegészül egy negyedik érzékelővel az ultraibolya (UV) tartományban. Ez a tény a színek minőségileg más jellegű megkülönböztetését teszi lehetővé. Hogy megtudjuk, miként látják egymást ezek a lepkék, a CIE színingermérő rendszerre alapozva háromdimenziós leképezést készítettünk (lásd [7], 5. függelék). A kapott adatokat ábrázolva (6. ábra és [7], 2. függelék) láthatjuk, ahogy az egyes lepkefajok jobban elkülönülnek, mint a korábban használt CIE diagram esetében [8].
Míg a kutatók a lepkeszárny fonák oldalának mintázatát használják a fajok azonosításra (ami laboratóriumi, preparált lepkék esetében jól működik), addig a repülő hím lepkéknek ezt a fajra jellemző apró mintázatot kellene felismerniük egy vizuálisan összetett növényi környezetben (harmincszorosan lassított videofelvétel: [7], 3. függelék). Hasonlóan, a nőstényeknek a gyors szárnymozgású hímeket kellene felismerniük (harmincszorosan lassított videofelvétel: [7], 1. függelék). Kérdéses, hogy a lepkék agya képes-e feldolgozni ilyen gyorsan a vizuális információkat (egy szárnycsapás 10-20 ms idejű) és a szárnyfonák bonyolult mintázatát sikeresen felismerni. Vegyük azt is figyelembe, hogy az egyes fajok szárnyfonákjának mintázata legtöbbször csak kis mértékben tér el; továbbá, a lepkék között gyors röptűnek számító boglárkalepkéknél a távoli felismerés nagyon előnyös lehet, hogy megelőzhessék a sikertelen párzási próbálkozásokat.
A tanulmányozott hímek ugyan rendelkeznek illatpikkelyekkel, de ezek szerepe a párválasztásban csak részben tisztázott, és az eredmények arra utalnak, hogy szerepük nem egyezik meg az éjszakai lepkéknél tapasztalt nagy távolságú, feromon alapú vonzással. A saját élőhelyükön történt megfigyelések alátámasztják, hogy a hímek és nőstények illatanyagainak is csak testközelből van szerepük. Nem ismert, hogy a vizsgált lepkefajok ivarszervi szerkezete önmagában mennyire hatékonyan gátolja a kereszteződéseket (hibridek létrejöttét), hiszen a fajok mesterséges körülmények között párosíthatók. Ha az ivarszervek szerkezete eléggé eltérő is lenne, a párzási próbálkozást általában egy hosszas udvarlás előzi meg. Így nagyon fontos lehet bármilyen optikai vagy kémiai jelzés, ami segíti a fajok megkülönböztetését, hiszen ezáltal felesleges energiabefektetés előzhető meg. Kísérletileg is bizonyították a kémiai jelzés hatását a Lycaeides argyrognomon lepkefajnál: a hím által kibocsátott illatanyagok hatására a nőstény a befogadó testhelyzetet veszi fel.
A boglárkákra vonatkozóan az irodalom hivatkozik arra, hogy ha a nőstény vonzza a hímet, akkor az közel repül és megkezdi az udvarlást. Ez ellentmond annak az egyértelmű ténynek, hogy a hím minden tekintetben sokkal feltűnőbb. Ha a hím a látványában (sokkal élénkebb szín), az illatanyagok tekintetében (jelentős mennyiségű illatpikkely jelenléte miatt intenzívebb kémiai jelzés, mint a nősténynél) és a mozgékonyságában is feltűnőbb, felmerülhet a kérdés, hogy mi a nőstény jelzése? Szintén ez a kérdés vetődik fel, ha a nőstény szárnyfonákján található sárga foltokat tekintjük a legfontosabb jelzésnek. Ezek a foltok a hímek számára abban az optikailag bonyolult környezetben, ami a fűben üldögélő nőstényt körülveszi, gyakorlatilag láthatatlanok. A kísérletek szerint a Polyommatus icarus szeme a szárnyak kék színére a legérzékenyebb.
Szabadtéri megfigyeléseink és mások munkái is azt bizonyítják, hogy a nőstények és hímek viselkedése nagyon eltérő. A bábból kikelt nőstények nem mutatkoznak, a növényzetben elbújva a kikelés helyén maradnak, és csak akkor repülnek, amikor a hím közeledik, hogy felhívják magukra a figyelmet. Bár szabad téren nem teszteltük, de az a hipotézisünk, hogy erre a hívó táncra csak akkor kerül sor, ha megfelelő színű hím közeledik. A jelenség nehezen figyelhető meg, mivel a nőstények leginkább párzás, táplálkozás vagy petézés közben láthatók.
A szubtrópusi-trópusi erdőkben található rokon boglárkafajok más stratégiát alkalmaznak: a nőstények még a bábban tartózkodva erős illatanyagokat bocsátanak ki, és ezzel vonzzák a hímeket. A mérsékelt klímájú füves területeken nincs ilyen megfigyelés. A vizsgált boglárkafajok hernyói hangyák járataiban telelnek, onnan is kelnek ki, és amikor elhagyják a talajt, egy fűszálon kapaszkodva tudják a szárnyaikat kifeszíteni és megszárítani. Közvetlenül ezután a párkeresés és a párzás is megtörténik, ezért a gyűjteményekben is nehéz nem megtermékenyített nőstényt találni.
Bár a pikkelyszerkezet teljes háromdimenziós szerkezetének feltárását leginkább a TEM tomográfia tenné lehetővé, SEM és ugyanolyan területről készített TEM képek megfelelően feldolgozva, elég sok jellemzőt megmutatnak. A cikkünkben vizsgált szárnypikkelyek első látásra nagyon hasonlóak, mégis jól érzékelhető az általuk keltett szín különbsége. A szerkezet MNH-elemzése megmutatta, hogy a pikkelyek adatai jellemzőek az egyes fajokra. Korábban az Albulina metallica faj esetében megmutattuk, hogy a szivacsos szerkezet parányi eltérései miképpen befolyásolják a szárnyak színét [3]. A fényvisszaverés spektrális maximumának legfontosabb meghatározója a szivacsszerű szerkezetben a lyukak távolsága és a kitin kitöltési tényezője. A Biophot Analyzer programmal meg lehet határozni felszíni (SEM képek) és mélységi (TEM képek) adatokat is. A tárgyalt fajok esetében 9 adatot határoztunk meg a fajok hatékony megkülönböztetéséhez (2. táblázat ). Megfelelően nagy számú bemeneti adat mellett automatikusan is el lehet különíteni az egyes fajokhoz tartozó mikroszkópos képeket. Az algoritmus által elkövetett legnagyobb hiba a 3. táblázatban a P. icarus faj P. thersites ként való azonosítása. Ezek visszaverési spektruma (3. ábra) nagyon hasonló, a spektrális adatokat összehasonlító MNH is itt követett el hibát (4. táblázat). A másik a P. coridon egyik példányának P. damonként való azonosítása. Ez a hiba a lepkék életkorának előrehaladásával a felső rétegben levő kék pikkelyek elvesztésének tulajdonítható. A megmaradó alappikkelyek barna melanin festékanyagot tartalmaznak, és ez okozza a fényvisszaverés növekedését a 600-800 nm hullámhossztartományon.
Ha összességében tekintjük a szerkezet és a szín MNH-elemzését, láthatjuk, hogy a fajra jellemző színek illeszthetőek a fajra jellemző szerkezetekkel. Tekintettel arra, hogy a szerkezet SEM képeken szabad szemmel láthatóan nagyon kis megváltozása felelős a jelentős színi eltérésekért, a fotonikus nanoarchitektúra feltehetően rendkívül hatékonyan és stabilan van beírva a fajok genetikai állományába. Nagyon valószínű, hogy a nem megfelelő színnel (és szerkezettel) rendelkező egyedek hamar kiküszöbölődnek a párzási folyamatból. Ez azt sugallja, hogy ahol a színnek szerepe van a kiválasztódásban, érdemes részletes szerkezeti és optikai elemzésnek alávetni a rokon fajok pikkelyszerkezeteit, mert egy ilyen munka számítógépes modellekkel kiegészítve új ötletekkel szolgálhat a bioinspirált fotonikus nanoszerkezetek tervezésében.
Figyelembe véve a lepkék szemében levő UV-érzékeny színérzékelő receptortípust is, a lepkék által érzékelt színtér egy dimenzióval kiegészül az emberi színlátáshoz képest. Így a 110 példány spektrumait egy 3D diagramon ábrázolhatjuk, ahol a pontok közötti kisebb távolságok a színek közötti kisebb különbségeket jelentenek, ahogy azt a lepkék látják. Következésképpen, az emberi szemhez illeszkedő 2D színingertérben részlegesen átfedő részek a 3D színingertérben jól elkülönülnek. Amint az elforgatott diagramon látszik ( [7], 5. függelék), vannak olyan metszetek, ahol bizonyos lepkefajok átfednek, mégis összességében a 3D-ben elkülönülnek. A negyedik színérzékelő lehetővé teszi az UV tartományon belüli jobb szétválasztást, és ez éppen egybeesik azzal a hullámhossztartománnyal, ahol a kék szárnyak spektrális jellegzetességei vannak. Az alábbiakban a 3D színingerdiagram és a repülésiidő-diagram együttes használatával megmutatjuk, hogy a különböző kék árnyalatú lepkék egy közös élettérben lehetnek anélkül, hogy zavarnák egymás párzását, míg a közel azonos árnyalatú lepkék az év más szakában repülnek.
Színüket tekintve ( 1. és 3. ábra) három csoportra oszthatjuk a vizsgált fajokat, és ez a feloszthatóság látható a 3D színingerdiagramon ( 6. ábra) is: lila (P. icarus, P. semiargus és P. thersites ), égszínkék (P. bellargus, P. daphnis és P. dorylas ) és zöldes (P. amandus, P. coridon és P. damon). A lila csoportban a visszaverési csúcs keskenyebb, mint a többi fajnál. Mivel a főmaximumok jól fedik egymást, szabad szemmel nehezen lehet megkülönböztetni a színüket ( 1. ábra). A másodlagos maximumoknak (250 nm körül) köszönhetően a 3D színingertérben jól elkülönülnek. A kék csoport spektruma kiszélesedik, a rövidebb hullámhosszú oldal helyzete nem változik, míg a hosszabb hullámhosszú lefutás jobbra tolódik. Egy plató is látható mindhárom spektrumon 325–400 nm között, ami egybeesik a lilák főmaximumának vállával. A 250 nm-nél látható másodlagos maximumok fedésben vannak. A zöldes csoportra a még erőteljesebb kiszélesedés jellemző. A P. amandus eltérő, amennyiben a visszaverési spektrumában egy határozott váll van 325 nm-nél, amint a lila csoportnál is láthattuk, és 250 nm körül van a másodlagos maximum. A zöldes csoport másik két tagja rövidebb hullámhosszaknál a kék csoport tagjaihoz hasonlít. Az 1. ábra SEM képein megfigyelhető, hogy a P. amandus szivacsos szerkezetének kitöltési tényezője nagyobb, mint a lila csoport tagjainak, míg a zöldes csoport másik két tagja, amelyeknek szintén nagyobb a kitöltési tényezője, mint a kék csoportban, ezzel együtt szélesebb szórást mutat a lyukak méretében, mint a kékek esetén. Modellezéssel kiegészített további munkára van szükség, hogy mélységében megértsük a szerkezet és a spektrum összefüggését. A tény, miszerint az MNH 91%-os találattal képes az egyes fajokhoz tartozó szerkezeteket azonosítani a visszaverési spektrum fajra jellemző kiegészítésével, arra utal, hogy a jól meghatározott szerkezeti jellegzetességek felelősek a szín kialakulásáért. Ez nem olyan egyszerű, mint a tökéletesen rendezett szerkezeteknél, ahol a törésmutatót állandónak véve, a szerkezet periodicitása egyértelműen meghatározza a fotonikus tiltott sáv helyét. Érdemes vizsgálni a szerkezetek és tulajdonságaik összefüggését, mivel a lepkék szárnyán található kvázirendezett szerkezetek bizonyítják ezen nanokompozitok lehetőségeit.
Az 5. ábrán a vizsgált lepkefajok eloszlását láthatjuk hónapok szerint. Egy, illetve két maximummal rendelkező csoportokat fedezhetünk fel, amelyek azt mutatják, hogy a P. icarus, P. dorylas és a P. bellargus esetében a nyár eleji nemzedék után egy második is repül a nyár vége felé. Bár nagy szórás van (a mintákat véletlenszerűen választottuk az 1930 és 2010 közötti időszakból), a hisztogramok határozott maximumot mutatnak. Hogy a kék szín a fajok megkülönböztetésére alkalmas távoli kommunikációs jel legyen, a színeknek nagy távolságból is jól láthatóan különbözniük kell. Ezért a megvizsgált boglárkák csoportjára jellemző negyedik fajta (UV-érzékeny) színérzékelő részletesebb megkülönböztetést tesz lehetővé, mint az emberi szem. A kiegészítő UV-érzékelő a 300–450 nm tartományon segíti a színfelbontást. Ez épp az a tartomány, ahol a szárny visszaverésében a másodlagos maximum és a főmaximum bal oldala van. Tudomásunk szerint első alkalommal végeztük el a számolást a CIE ábrázolást felhasználva a négy érzékenységi görbére a lepkék látásának vizsgálatára (6. ábra). Ez a megközelítés hasznos lehet más lepkék vizsgálatának esetében is. Fontos viszont megjegyezni, hogy bizonyos fajok akár nyolcféle színérzékelővel is rendelkezhetnek, így ezekhez többdimenziós ábrázolás szükséges.
A repülési időket tekintve, a lila csoportban a P. icarus és a P. thersites nagyon hasonló színű, de a P. thersites rajzása (egy nemzedék) beékelődik a két P. icarus nemzedék közé. A P. thersites repülési idejének eltolódása július vége felé megengedett, mivel az akkori példányok már öregek, ezáltal kopottak, és nem zavarják a P. icarus második generációjának nászrepülését. Átfedés figyelhető meg a P. icarus és a P. semiargus esetében is, de a hisztogramon láthatjuk, hogy a P. icarus első nemzedéke május közepén-végén tetőzik, amikor az első P. semiargusok megjelennek, és egészen hosszan tart a repülési idejük, június közepi tetőzéssel. A P. thersites példányok csak a P. semiargus nemzedék végén kezdenek repülni, amikor már csak idős példányok fordulnak elő.
Az égszínkék csoportból a P. bellargus és a P. dorylas is kétnemzedékes. A P. bellargus rendelkezik a legkorábbi és legkésőbbi tetőzéssel, és bár van valamennyi átfedés a P. dorylas repülésével, spektrálisan jól láthatóan különböznek. A P. dorylas második nemzedékének repülési ideje teljesen átfed a P. daphnis előfordulásával. Bár a színük lehetővé teszi a megfelelő megkülönböztetést, a két faj elkülönülését azon egyedi jelenség is segíti, hogy a P. daphnis nőstényei mindig kékek.
A zöldes csoport tagjai egynemzedékesek. Elsőként a P. amandus jelenik meg, ami színében is eltér a többitől. Június közepére már megfakul a szárny színe annyira, hogy ne zavarja a megjelenő P. damon faj párzását. A P. coridon és a P. damon váltják egymást, a rendszerint nagy számban kirajzó P. coridon már csak az idős, kevés számú P. damon hímmel találkozhat az élőhelyen.
A fenti csoportosulásokon kívül meg kell jegyezzük a kétnemzedékes P. icarus, P. dorylas és P. bellargus esetét. Május közepén vagy végén tetőzik az első nemzedékük, de a színük jellegzetesen elkülönül, tehát ennek alapján az ugyanazon fajhoz tartozó egyedek azonosítani tudják egymást.
Kilenc közeli rokon (és azonos földrajzi helyen élő) boglárka lepkefaj részletes szerkezeti és spektrális vizsgálata megmutatta, hogy a hímek szárnyának kék színe és a színért felelős fotonikus nanoszerkezet fajra jellemző. Mesterséges neurális hálózatot alkalmazva, a szerkezeti vagy a spektrális adatok alapján is a fajokat 90% feletti találati aránnyal lehet azonosítani. A fajok repülési idejét tekintve az látszik, hogy a hasonló színű lepkék időben elkülönülnek. A lepkék ultraibolya-érzékeny fotoreceptor-típusát figyelembe vevő, háromdimenziós színingertérben ábrázolva, az eltérő fajok szárnyszínei jobban elkülönülnek egymástól, mint az emberi látáson alapuló, kétdimenziós színingertérben. A repülési időszakokat tekintve, a vizsgált 9 boglárkafaj kék színe lehetővé teszi a biztonságos párkeresést, illetve a versenytársak eredményes kiszűrését.
A szerkezet-szín összefüggés vizsgálata modellezéssel együtt lehetővé teszi új utak feltárását a bioinspirált, kívánt színárnyalatú mesterséges fotonikus nanoarchitektúrák tervezésében és azok gyakorlati alkalmazásában.
_______________________
A munka az OTKA PD 83483, és a Bolyai János Kutatási Ösztöndíj támogatásával készült.